

The mTORC1/4E-BP1 axis represents a critical signaling node during fibrogenesis
- PMID: 30602778
- PMCID: PMC6315032
- DOI: 10.1038/s41467-018-07858-8
The mTORC1/4E-BP1 axis represents a critical signaling node during fibrogenesis
Erratum in
-
Author Correction: The mTORC1/4E-BP1 axis represents a critical signaling node during fibrogenesis.Nat Commun. 2020 Sep 11;11(1):4680. doi: 10.1038/s41467-020-18621-3. Nat Commun. 2020. PMID: 32917879 Free PMC article.
Abstract
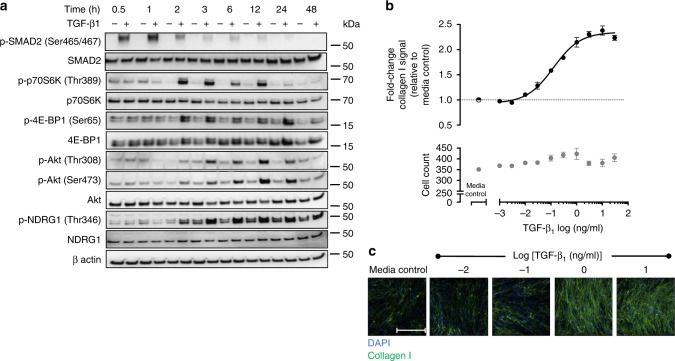
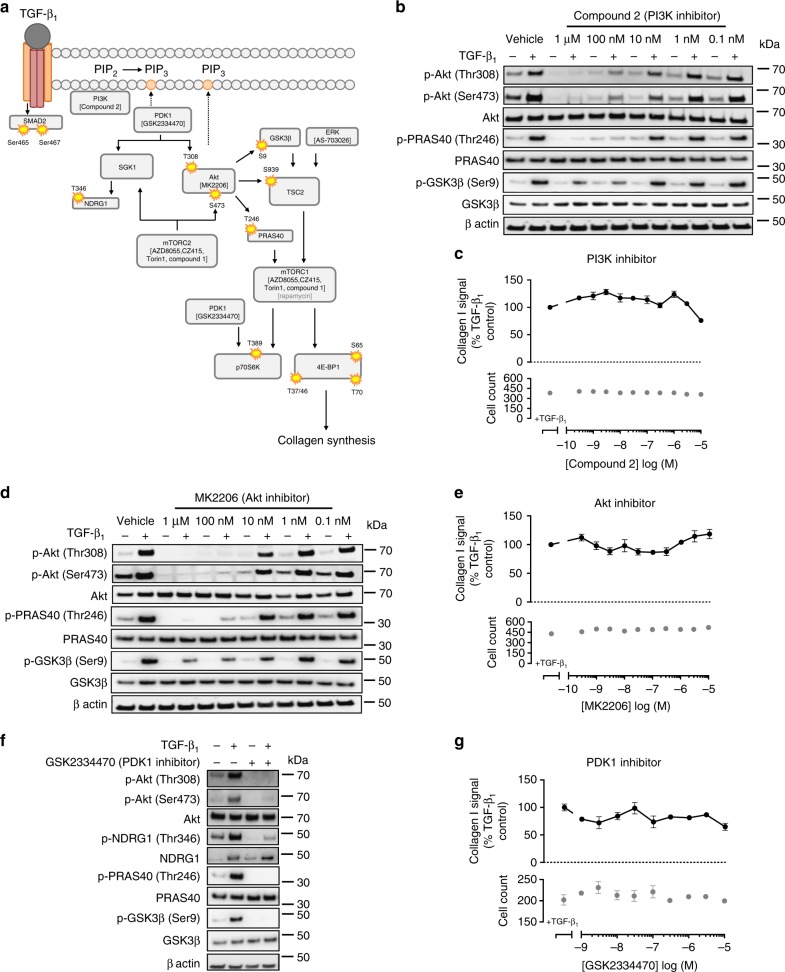
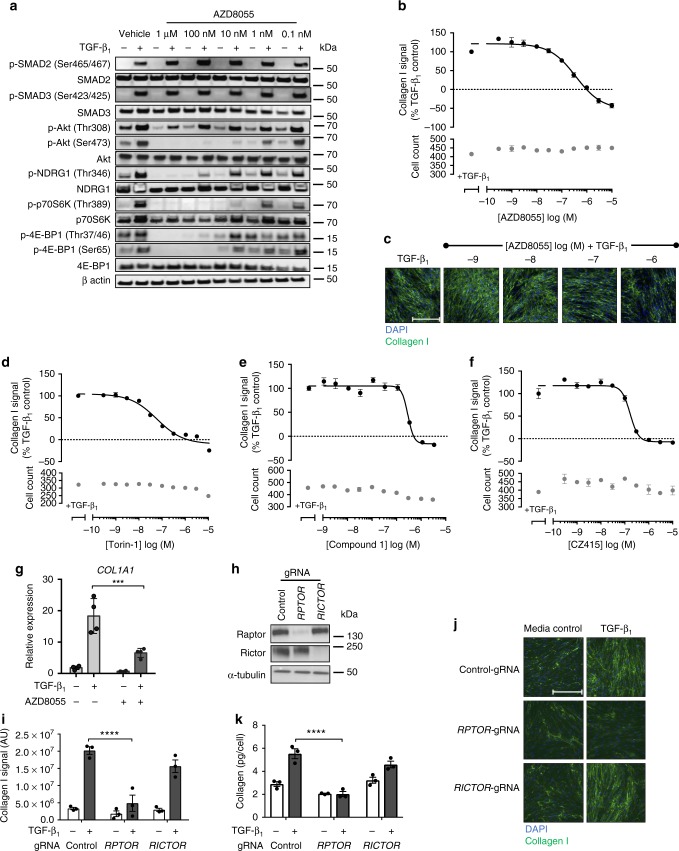
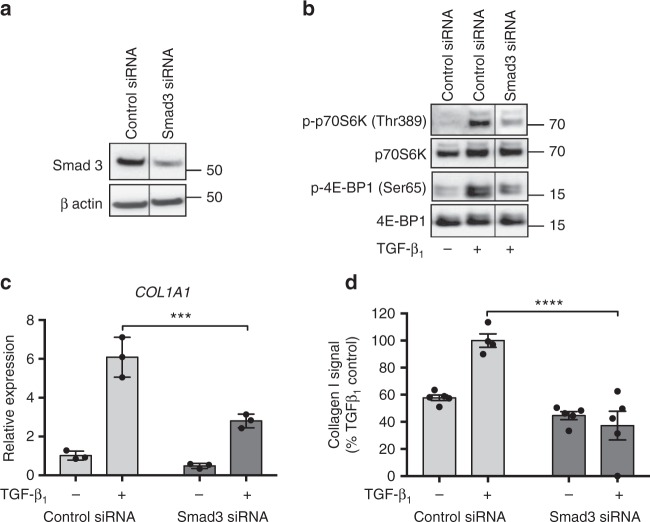
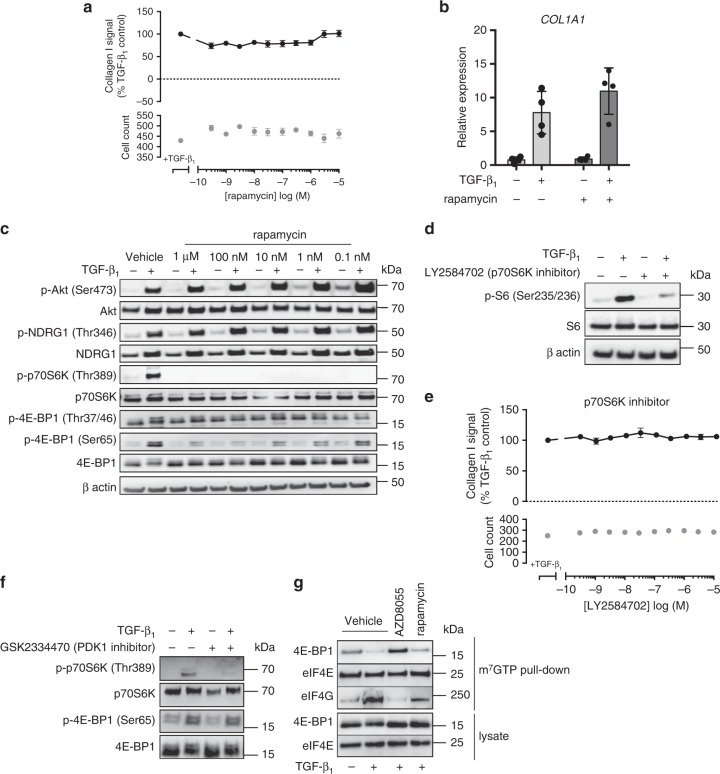
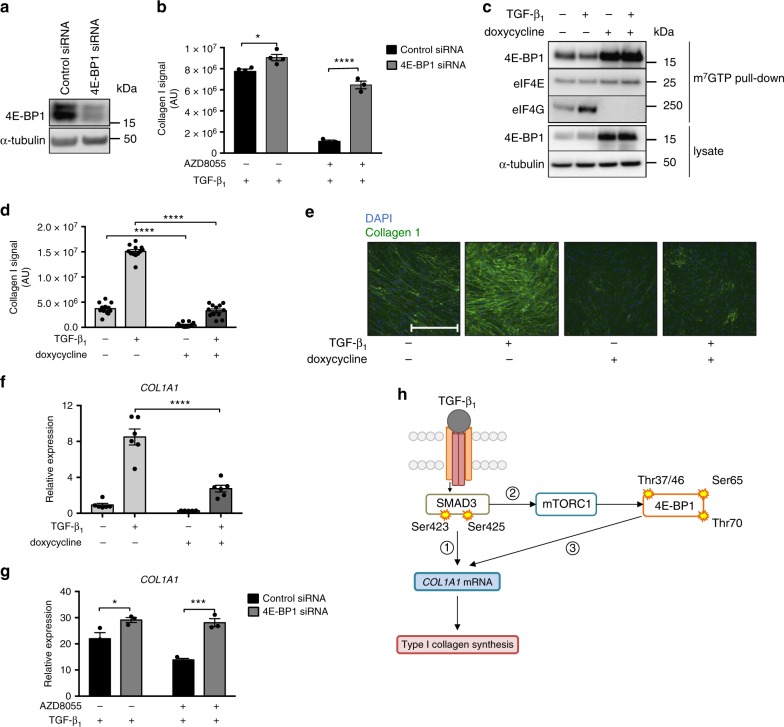
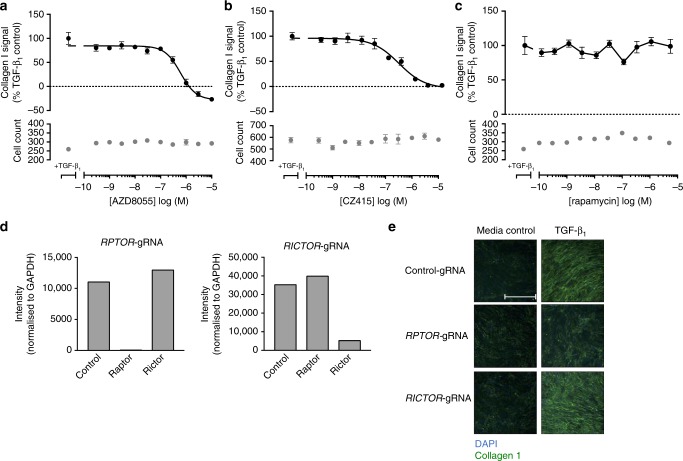
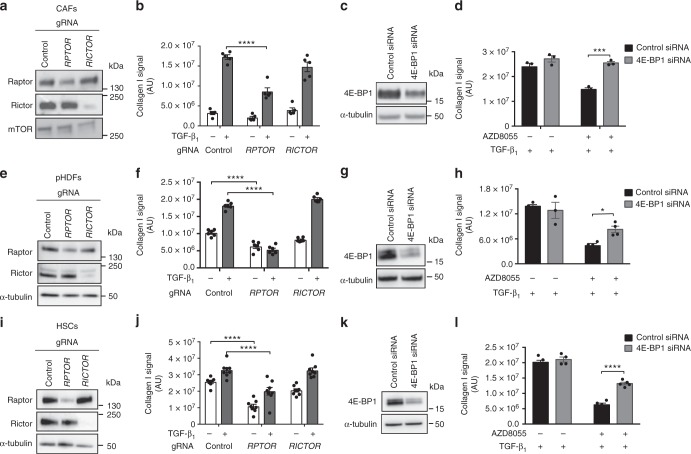
Myofibroblasts are the key effector cells responsible for excessive extracellular matrix deposition in multiple fibrotic conditions, including idiopathic pulmonary fibrosis (IPF). The PI3K/Akt/mTOR axis has been implicated in fibrosis, with pan-PI3K/mTOR inhibition currently under clinical evaluation in IPF. Here we demonstrate that rapamycin-insensitive mTORC1 signaling via 4E-BP1 is a critical pathway for TGF-β1 stimulated collagen synthesis in human lung fibroblasts, whereas canonical PI3K/Akt signaling is not required. The importance of mTORC1 signaling was confirmed by CRISPR-Cas9 gene editing in normal and IPF fibroblasts, as well as in lung cancer-associated fibroblasts, dermal fibroblasts and hepatic stellate cells. The inhibitory effect of ATP-competitive mTOR inhibition extended to other matrisome proteins implicated in the development of fibrosis and human disease relevance was demonstrated in live precision-cut IPF lung slices. Our data demonstrate that the mTORC1/4E-BP1 axis represents a critical signaling node during fibrogenesis with potential implications for the development of novel anti-fibrotic strategies.
Conflict of interest statement
This work was part-funded by a collaborative framework agreement awarded to R.C.C. by GlaxoSmithKline. In terms of potential conflict of interest, the following authors: C.B.N., M.M., S.P., G.J., D.P., R.B.G., A.R.T., G.B., A.D.B., N.Z., and R.P.M. were employees of GlaxoSmithKline which has R&D programs for IPF at the time of writing. M.K. is an employee of Nordic Biosciences with a commercial interest in proprietary assay development for biomarker discovery. T.M.M. has no declarations directly related to this manuscript. He has, however, received industry-academic research funding from GlaxoSmithKline R&D, UCB, and Novartis and has received consultancy or speakers fees from Pellis, Astra Zeneca, Bayer, Biogen Idec, Boehringer Ingelheim, Cipla, Galapagos, GlaxoSmithKline R&D, ProMetic, Roche (and previously InterMune), Sanofi-Aventis, and UCB. All remaining authors declare no competing interests. All data presented have been recorded in full compliance with GSK data integrity.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
