

Genomic encoding of transcriptional burst kinetics
- PMID: 30602787
- PMCID: PMC7610481
- DOI: 10.1038/s41586-018-0836-1
Genomic encoding of transcriptional burst kinetics
Abstract
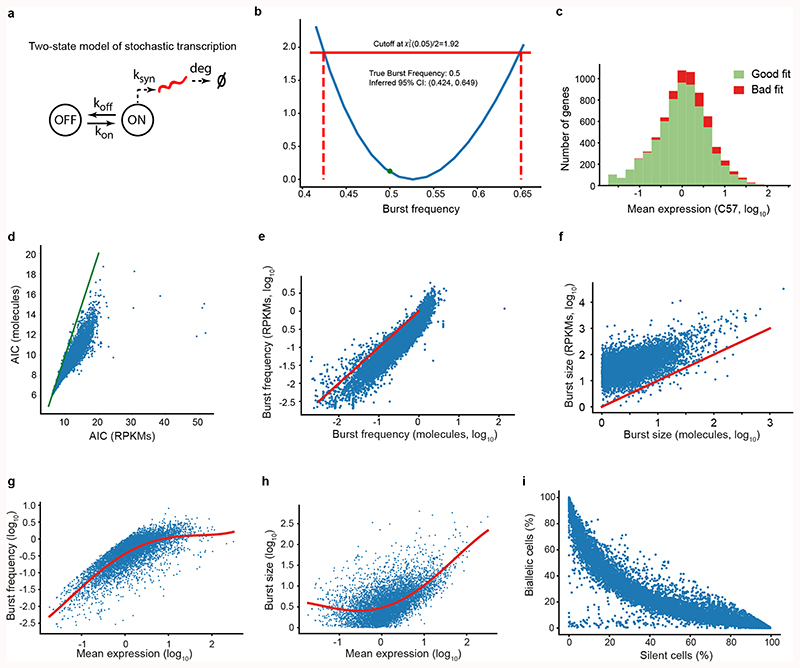
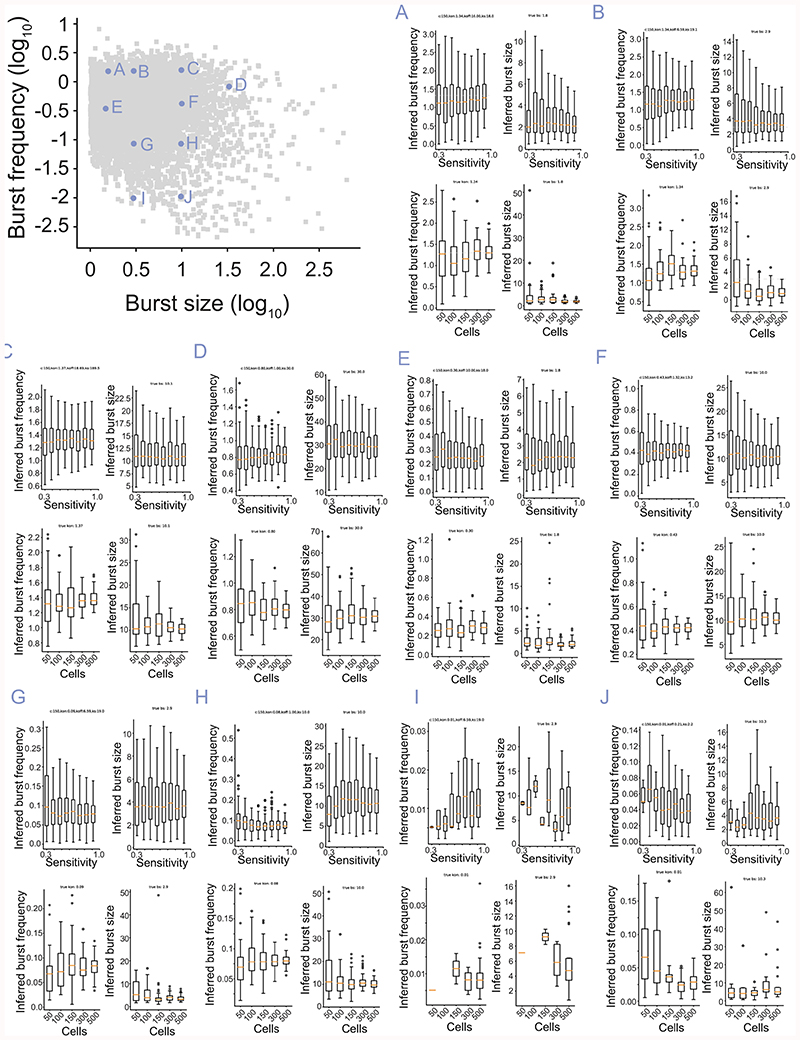
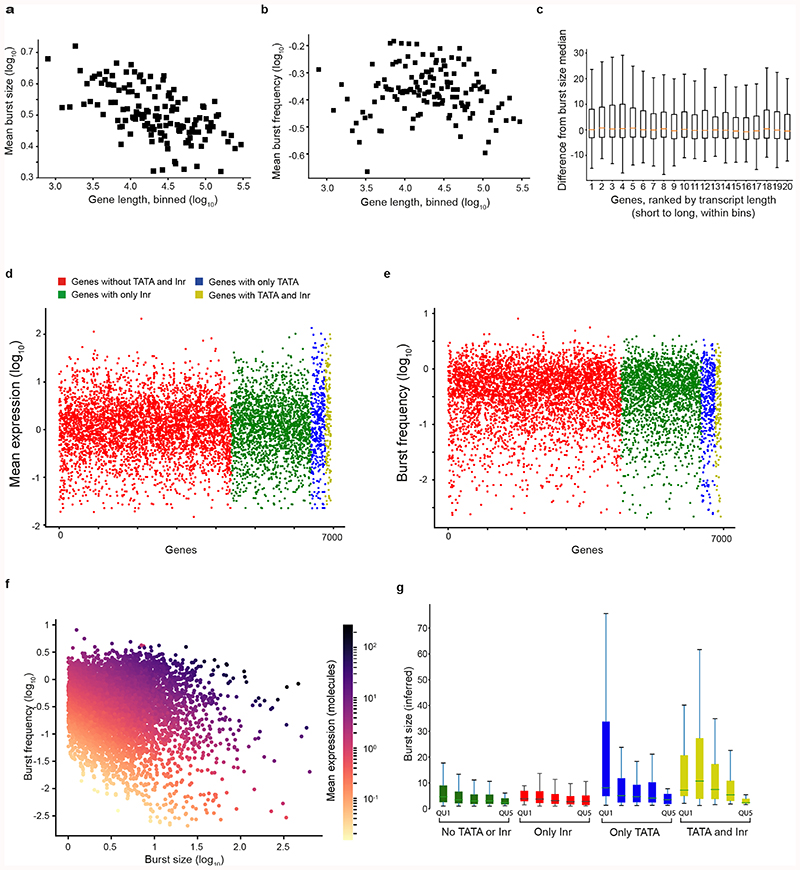
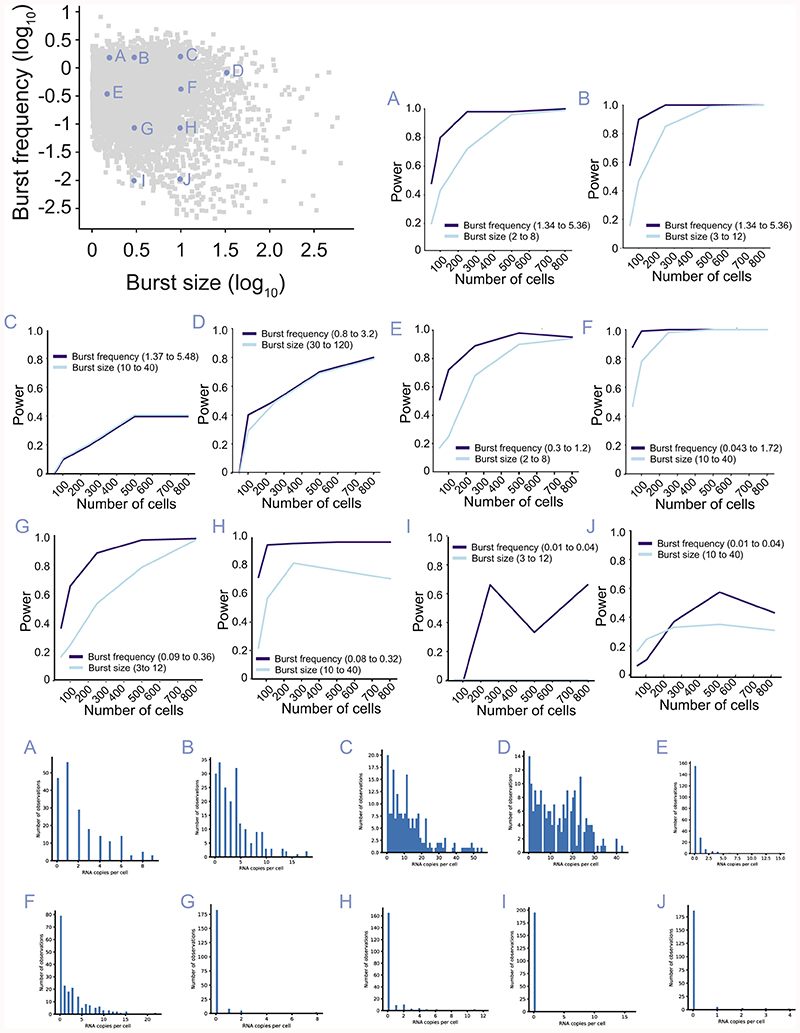
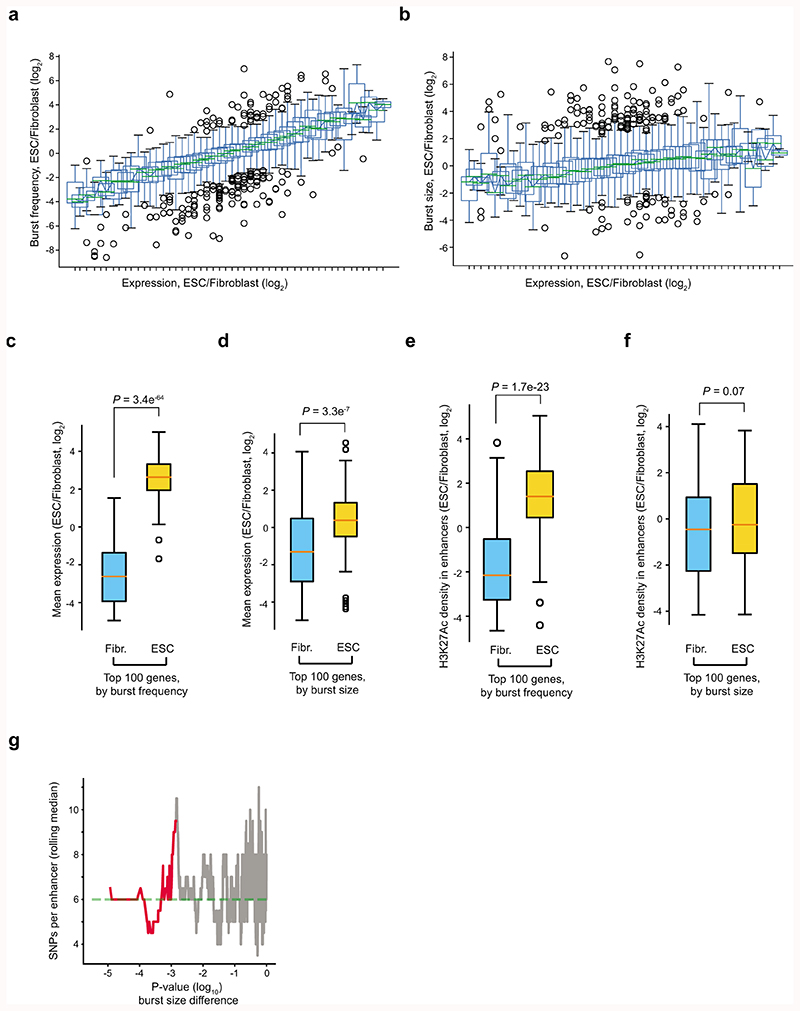
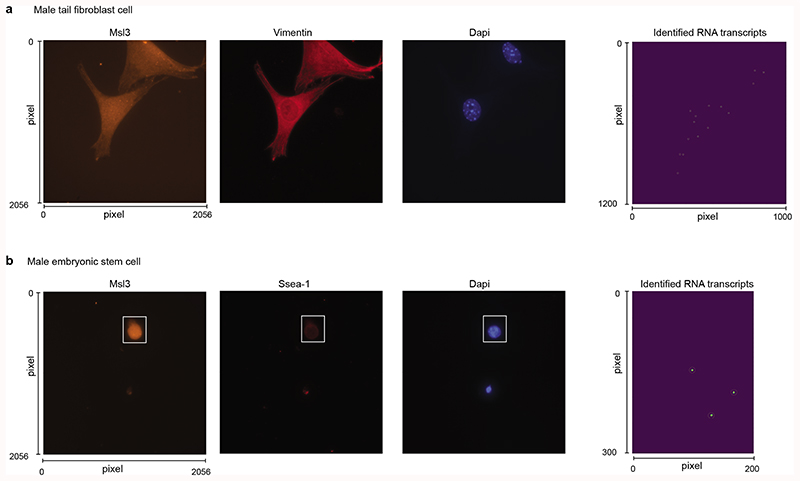
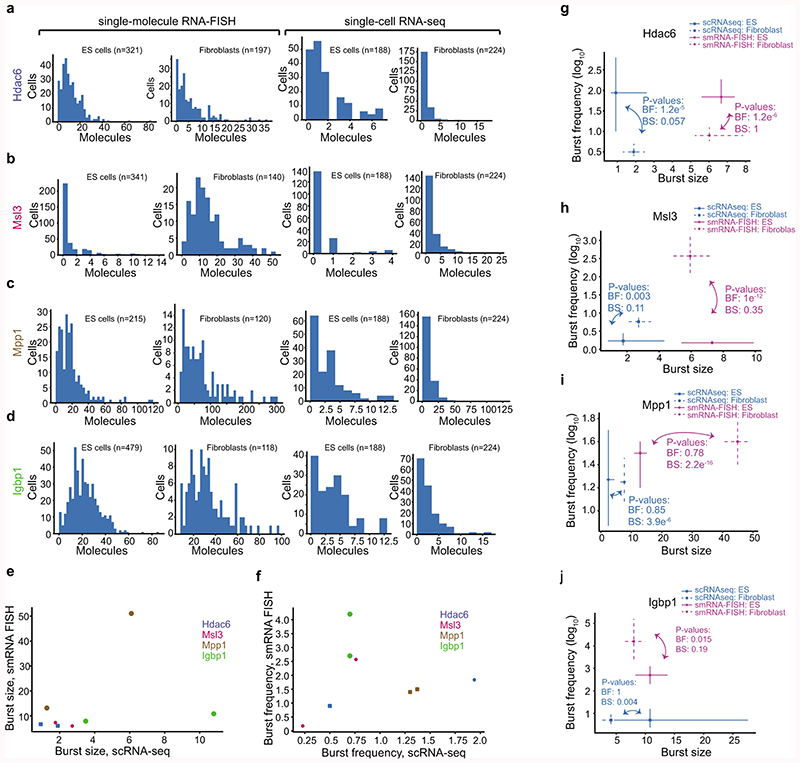
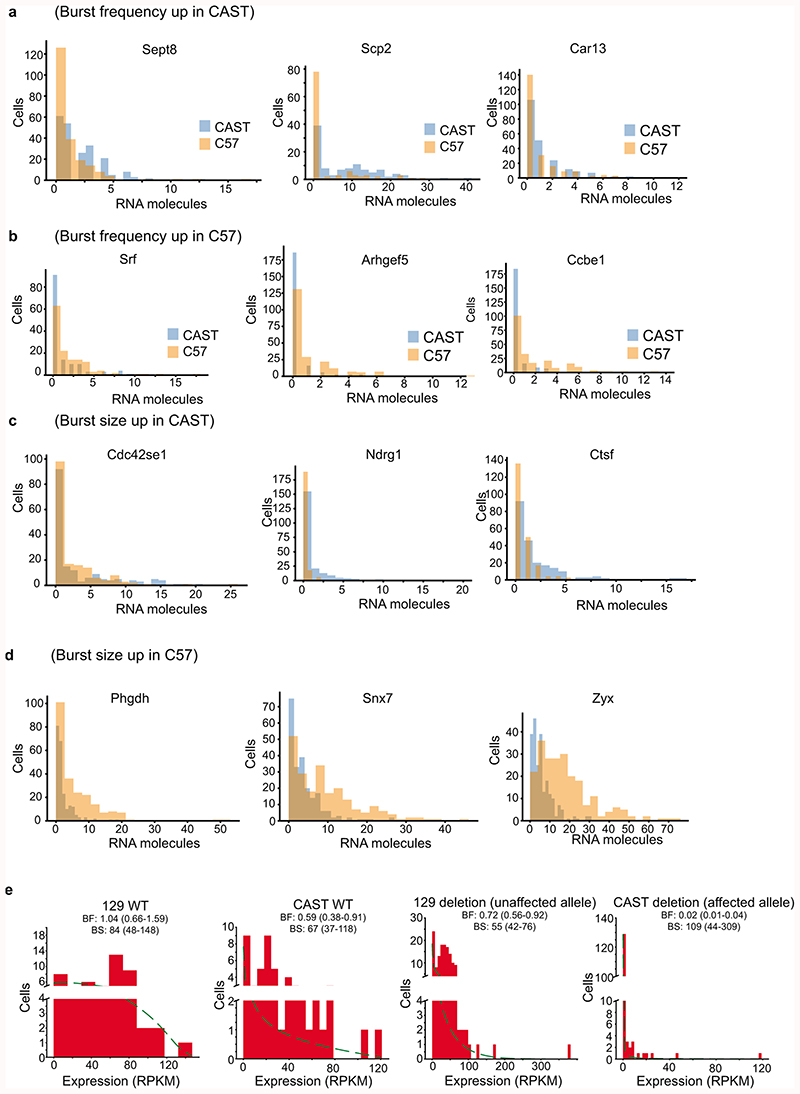
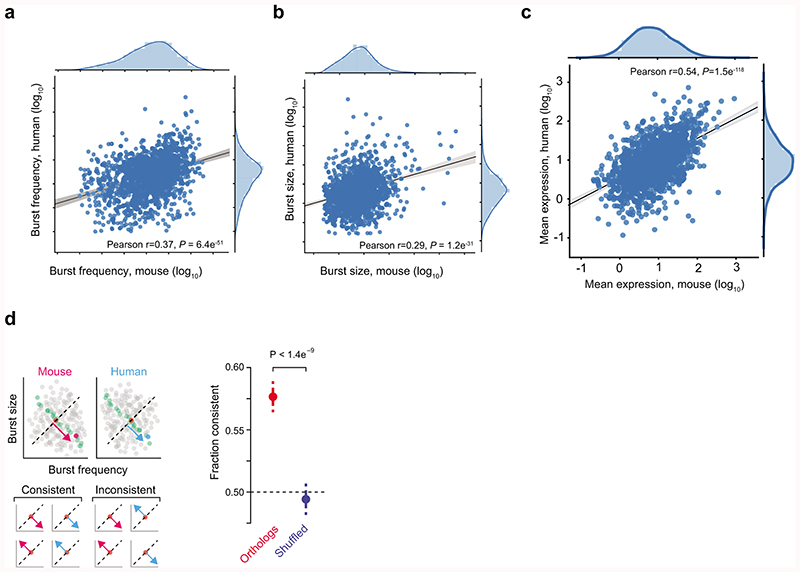
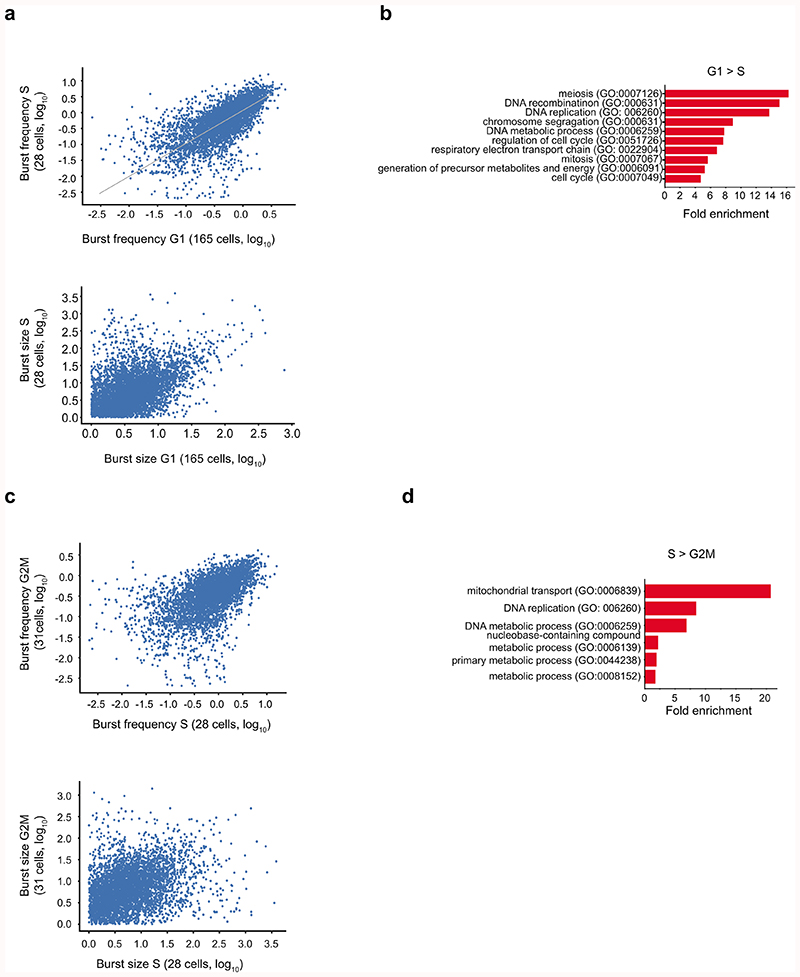
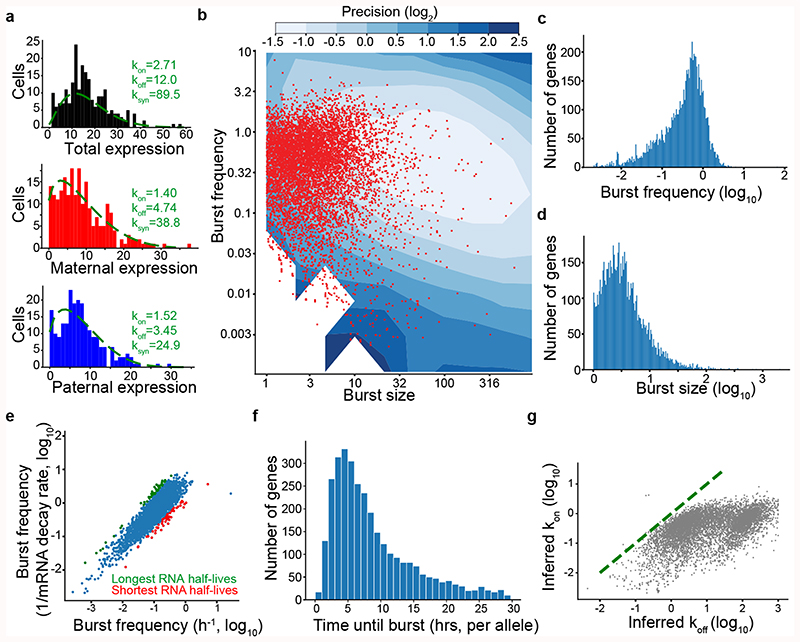
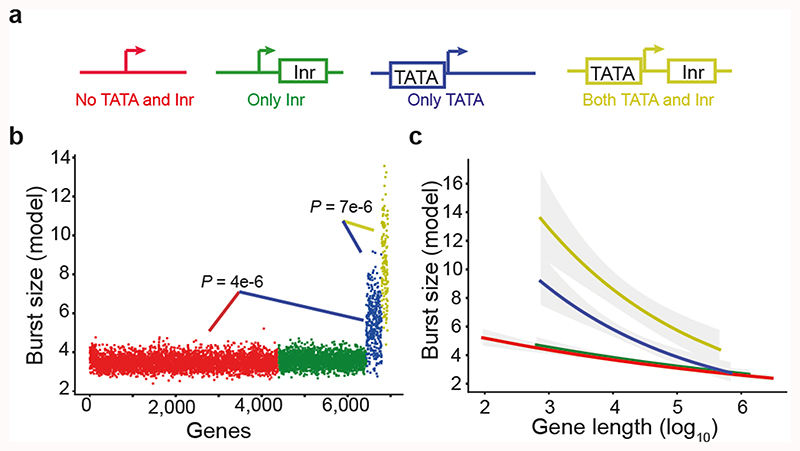
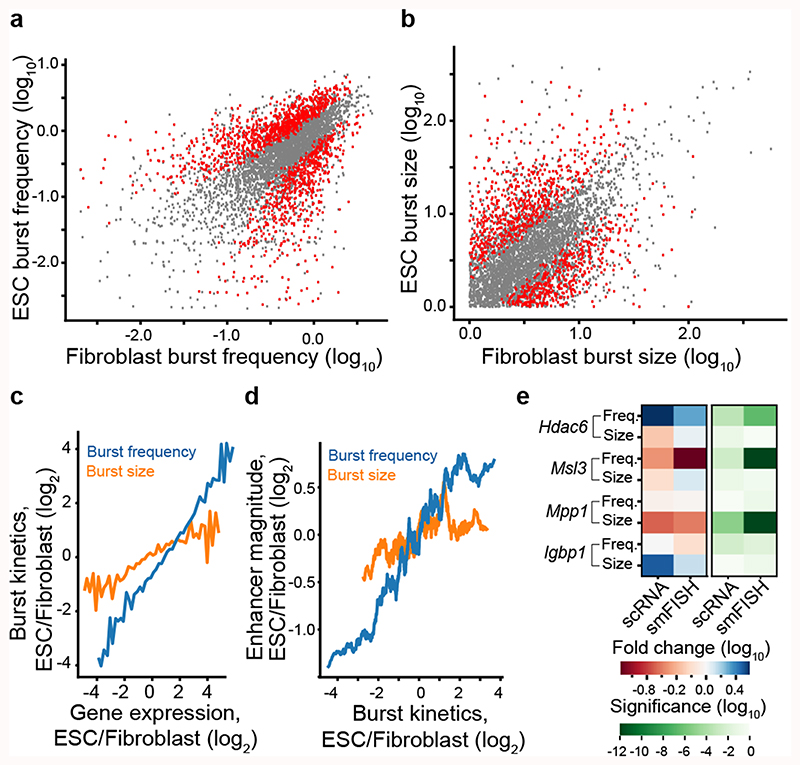
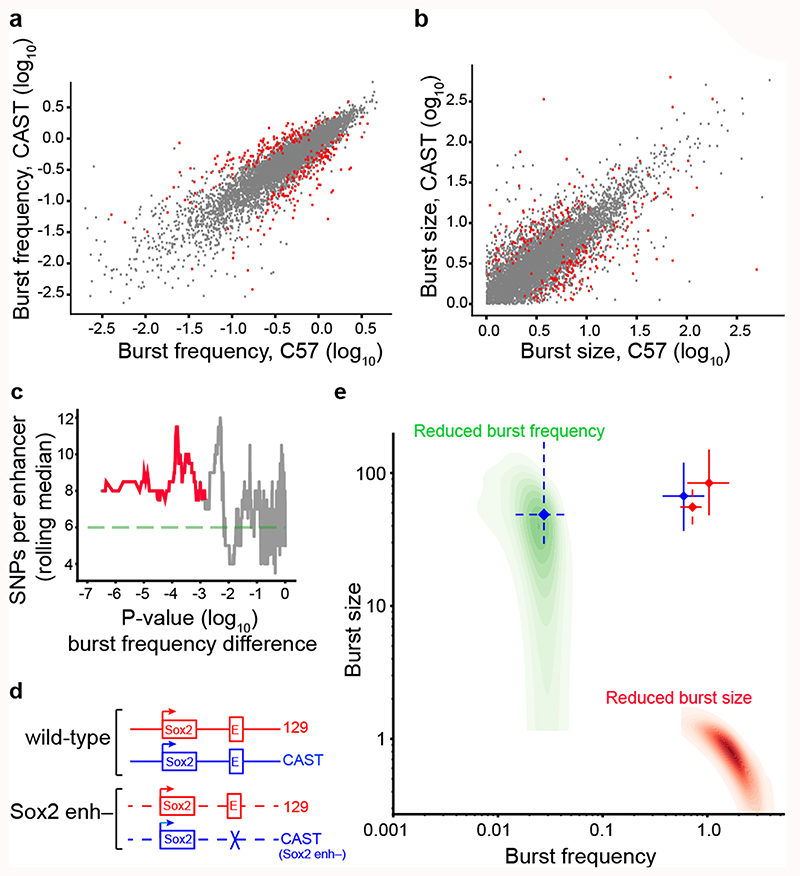
Mammalian gene expression is inherently stochastic1,2, and results in discrete bursts of RNA molecules that are synthesized from each allele3-7. Although transcription is known to be regulated by promoters and enhancers, it is unclear how cis-regulatory sequences encode transcriptional burst kinetics. Characterization of transcriptional bursting, including the burst size and frequency, has mainly relied on live-cell4,6,8 or single-molecule RNA fluorescence in situ hybridization3,5,8,9 recordings of selected loci. Here we determine transcriptome-wide burst frequencies and sizes for endogenous mouse and human genes using allele-sensitive single-cell RNA sequencing. We show that core promoter elements affect burst size and uncover synergistic effects between TATA and initiator elements, which were masked at mean expression levels. Notably, we provide transcriptome-wide evidence that enhancers control burst frequencies, and demonstrate that cell-type-specific gene expression is primarily shaped by changes in burst frequencies. Together, our data show that burst frequency is primarily encoded in enhancers and burst size in core promoters, and that allelic single-cell RNA sequencing is a powerful model for investigating transcriptional kinetics.
Conflict of interest statement
Figures
Comment in
-
Enhancers and promoters regulate burst kinetics.Nat Rev Mol Cell Biol. 2019 Mar;20(3):134-135. doi: 10.1038/s41580-019-0100-z. Nat Rev Mol Cell Biol. 2019. PMID: 30647472 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
