

C9orf72 arginine-rich dipeptide proteins interact with ribosomal proteins in vivo to induce a toxic translational arrest that is rescued by eIF1A
- PMID: 30604225
- PMCID: PMC6514073
- DOI: 10.1007/s00401-018-1946-4
C9orf72 arginine-rich dipeptide proteins interact with ribosomal proteins in vivo to induce a toxic translational arrest that is rescued by eIF1A
Abstract
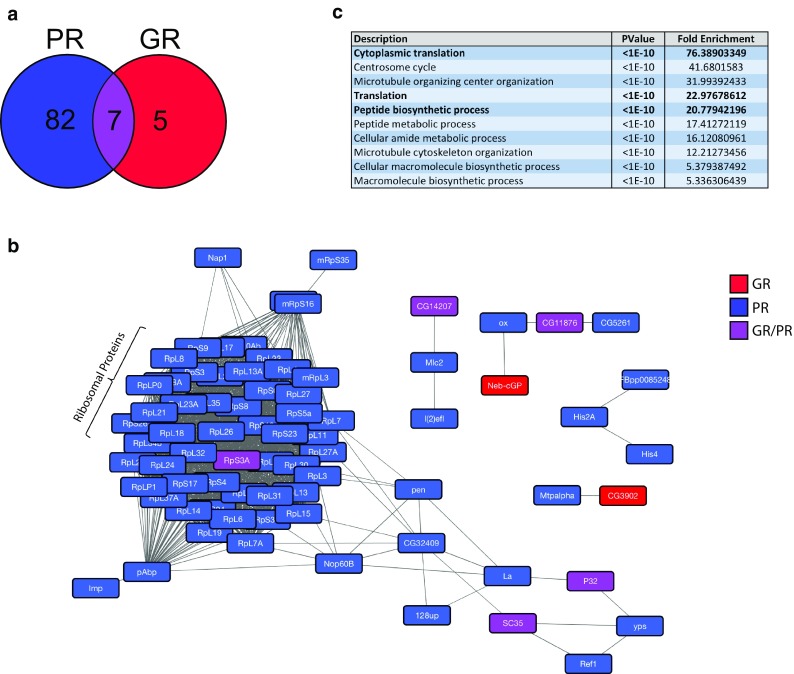
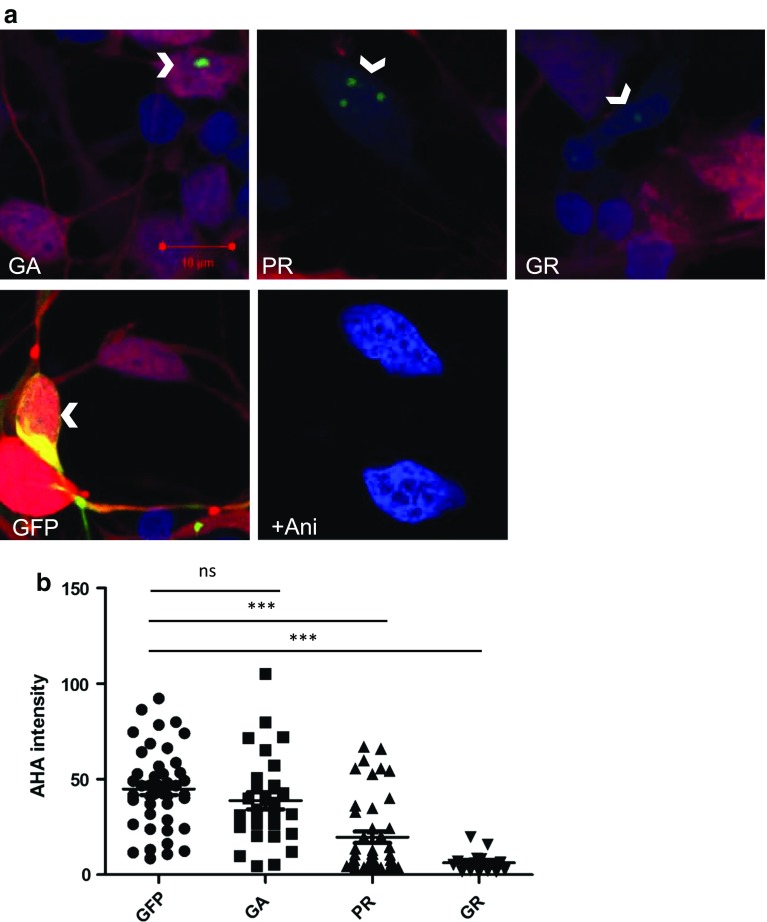
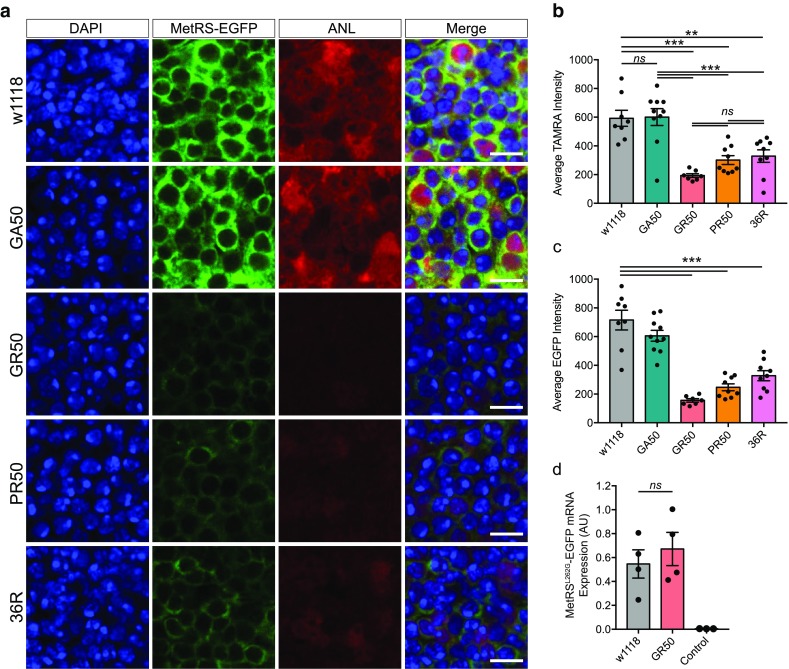
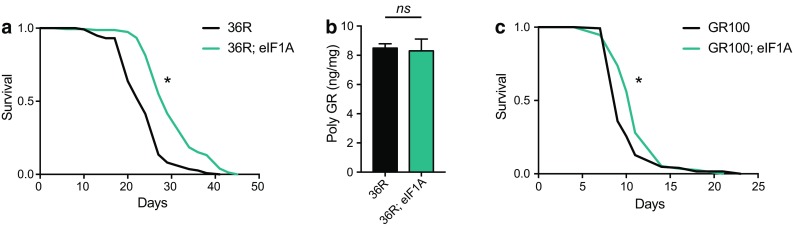
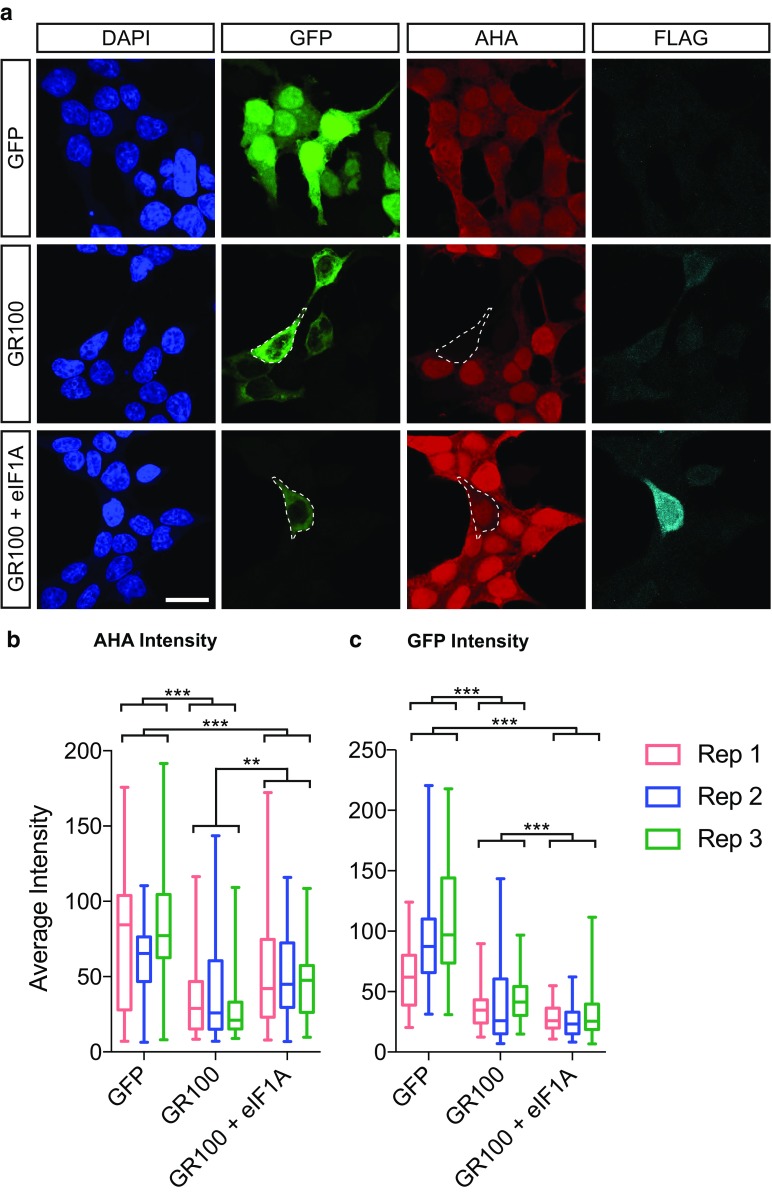
A GGGGCC hexanucleotide repeat expansion within the C9orf72 gene is the most common genetic cause of both amyotrophic lateral sclerosis and frontotemporal dementia. Sense and antisense repeat-containing transcripts undergo repeat-associated non-AUG-initiated translation to produce five dipeptide proteins (DPRs). The polyGR and polyPR DPRs are extremely toxic when expressed in Drosophila neurons. To determine the mechanism that mediates this toxicity, we purified DPRs from the Drosophila brain and used mass spectrometry to identify the in vivo neuronal DPR interactome. PolyGR and polyPR interact with ribosomal proteins, and inhibit translation in both human iPSC-derived motor neurons, and adult Drosophila neurons. We next performed a screen of 81 translation-associated proteins in GGGGCC repeat-expressing Drosophila to determine whether this translational repression can be overcome and if this impacts neurodegeneration. Expression of the translation initiation factor eIF1A uniquely rescued DPR-induced toxicity in vivo, indicating that restoring translation is a potential therapeutic strategy. These data directly implicate translational repression in C9orf72 repeat-induced neurodegeneration and identify eIF1A as a novel modifier of C9orf72 repeat toxicity.
Keywords: ALS; C9orf72; Dipeptide; Drosophila; FTD.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- ERC-2017-COG 770244/ERC_/European Research Council/International
- FRATTA/JAN15/946-795/MNDA_/Motor Neurone Disease Association/United Kingdom
- 648716 - C9ND/ERC_/European Research Council/International
- WT_/Wellcome Trust/United Kingdom
- ISAACS/APR13/818-791/MNDA_/Motor Neurone Disease Association/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
