

A switch in the poly(dC)/RmlB complex regulates bacterial persister formation
- PMID: 30604752
- PMCID: PMC6318315
- DOI: 10.1038/s41467-018-07861-z
A switch in the poly(dC)/RmlB complex regulates bacterial persister formation
Retraction in
-
Retraction Note: A switch in the poly(dC)/RmlB complex regulates bacterial persister formation.Nat Commun. 2019 Jul 11;10(1):3119. doi: 10.1038/s41467-019-11189-7. Nat Commun. 2019. PMID: 31296865 Free PMC article.
Abstract
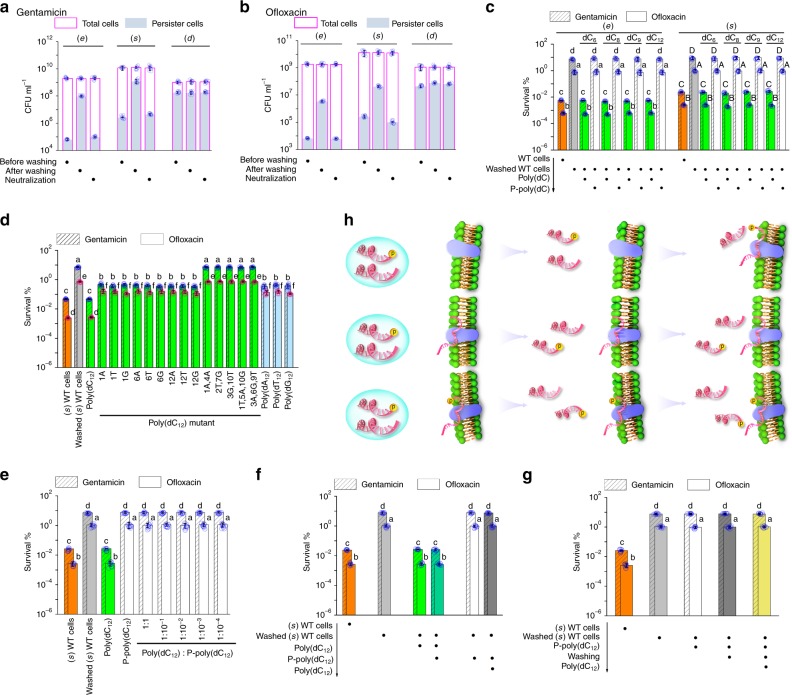
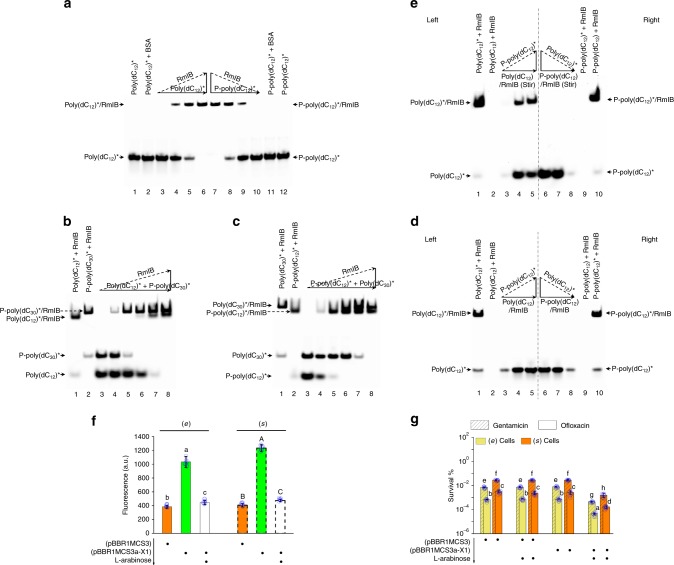
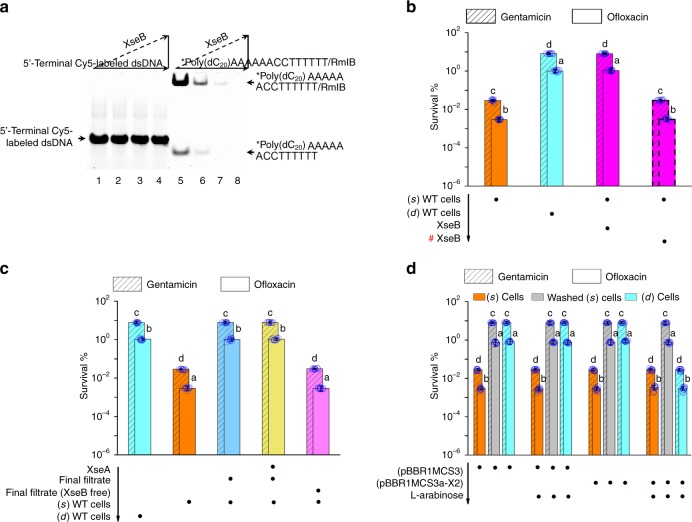
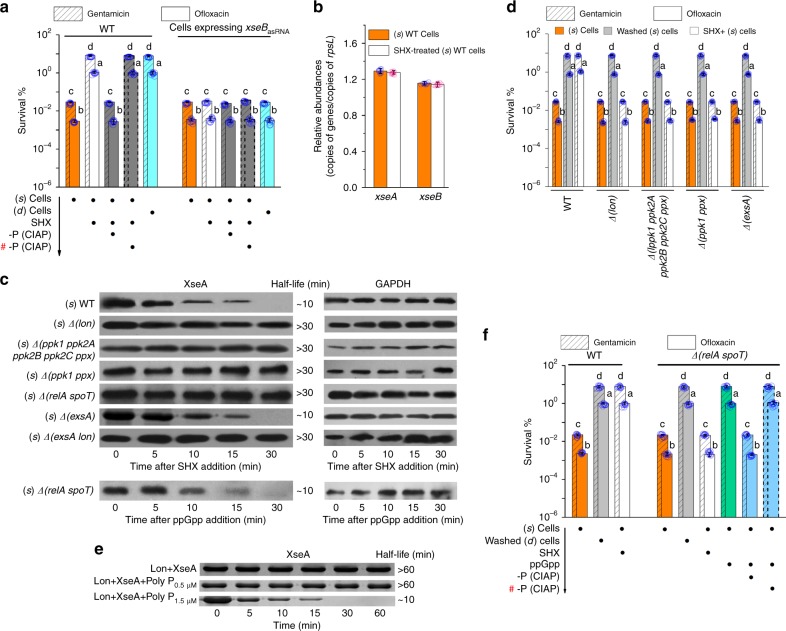
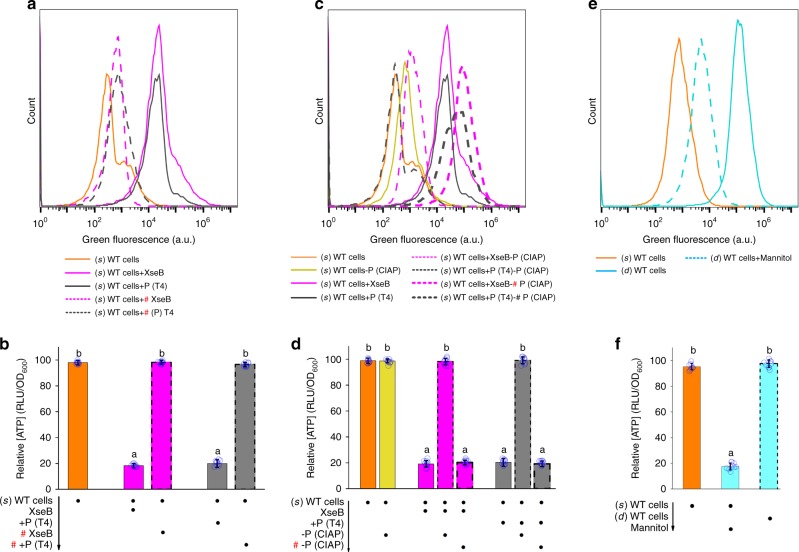
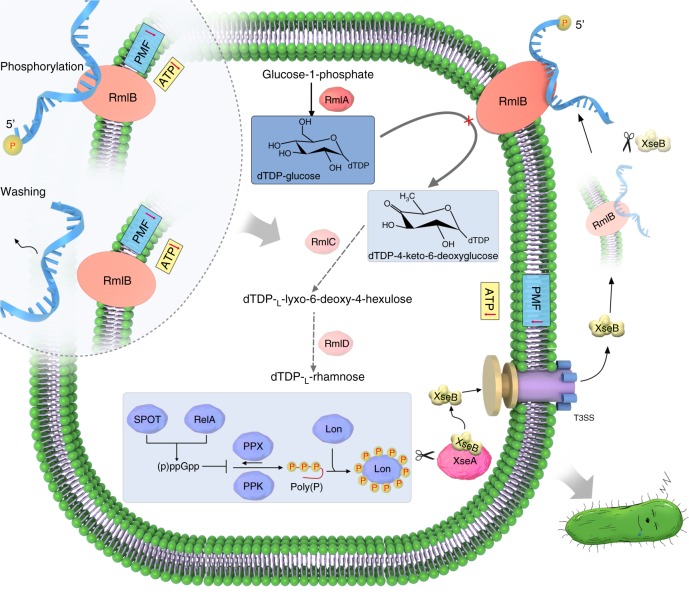
Bacterial persisters are phenotypic variants that tolerate exposure to lethal antibiotics. These dormant cells are responsible for chronic and recurrent infections. Multiple mechanisms have been linked to persister formation. Here, we report that a complex, consisting of an extracellular poly(dC) and its membrane-associated binding protein RmlB, appears to be associated with persistence of the opportunistic pathogen Pseudomonas aeruginosa. Environmental stimuli triggers a switch in the complex physiological state (from poly(dC)/RmlB to P-poly(dC)/RmlB or RmlB). In response to the switch, bacteria decrease proton motive force and intracellular ATP levels, forming dormant cells. This alteration in complex status is linked to a (p)ppGpp-controlled signaling pathway that includes inorganic polyphosphate, Lon protease, exonuclease VII (XseA/XseB), and the type III secretion system. The persistence might be also an adaptive response to the lethal action of the dTDP-L-rhamnose pathway shutdown, which occurs due to switching of poly(dC)/RmlB.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
