

IgA tetramerization improves target breadth but not peak potency of functionality of anti-influenza virus broadly neutralizing antibody
- PMID: 30605488
- PMCID: PMC6317788
- DOI: 10.1371/journal.ppat.1007427
IgA tetramerization improves target breadth but not peak potency of functionality of anti-influenza virus broadly neutralizing antibody
Abstract
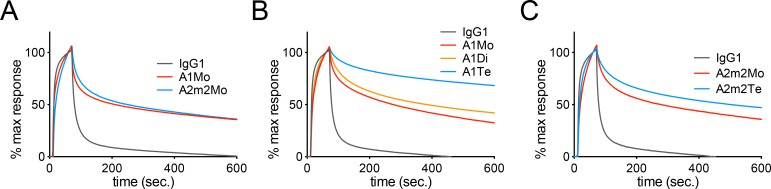
Mucosal immunoglobulins comprise mainly secretory IgA antibodies (SIgAs), which are the major contributor to pathogen-specific immune responses in mucosal tissues. These SIgAs are highly heterogeneous in terms of their quaternary structure. A recent report shows that the polymerization status of SIgA defines their functionality in the human upper respiratory mucosa. Higher order polymerization of SIgA (i.e., tetramers) leads to a marked increase in neutralizing activity against influenza viruses. However, the precise molecular mechanisms underlying the effects of SIgA polymerization remain elusive. Here, we developed a method for generating recombinant tetrameric monoclonal SIgAs. We then compared the anti-viral activities of these tetrameric SIgAs, which possessed variable regions identical to that of a broadly neutralizing anti-influenza antibody F045-092 against influenza A viruses, with that of monomeric IgG or IgA. The tetrameric SIgA showed anti-viral inhibitory activity superior to that of other forms only when the antibody exhibits low-affinity binding to the target. By contrast, SIgA tetramerization did not substantially modify anti-viral activity against targets with high-affinity binding. Taken together, the data suggest that tetramerization of SIgA improved target breadth, but not peak potency of antiviral functions of the broadly neutralizing anti-influenza antibody. This phenomenon presumably represents one of the mechanisms by which SIgAs present in human respiratory mucosa prevent infection by antigen-drifted influenza viruses. Understanding the mechanisms involved in cross neutralization of viruses by SIgAs might facilitate the development of vaccine strategies against viral infection of mucosal tissues.
Conflict of interest statement
National Institute of Infectious Diseases and Nippi, Inc. have filed patents regarding construction and use of tetrameric SIgA (application no. PCT/JP2015/070742). SS, TS, AA, YT, TU, KOG and HH are named as inventors on the application. YT, TU, and KOG are employees of Nippi, Inc.
Figures




Similar articles
-
Potential Role of Nonneutralizing IgA Antibodies in Cross-Protective Immunity against Influenza A Viruses of Multiple Hemagglutinin Subtypes.J Virol. 2020 Jun 1;94(12):e00408-20. doi: 10.1128/JVI.00408-20. Print 2020 Jun 1. J Virol. 2020. PMID: 32269119 Free PMC article.
-
IgA polymerization contributes to efficient virus neutralization on human upper respiratory mucosa after intranasal inactivated influenza vaccine administration.Hum Vaccin Immunother. 2018 Jun 3;14(6):1351-1361. doi: 10.1080/21645515.2018.1438791. Epub 2018 Apr 9. Hum Vaccin Immunother. 2018. PMID: 29425074 Free PMC article.
-
Broadly neutralizing anti-influenza virus antibodies: enhancement of neutralizing potency in polyclonal mixtures and IgA backbones.J Virol. 2015 Apr;89(7):3610-8. doi: 10.1128/JVI.03099-14. Epub 2015 Jan 14. J Virol. 2015. PMID: 25589655 Free PMC article.
-
Broadly neutralizing antibodies against influenza viruses.Antiviral Res. 2013 Jun;98(3):476-83. doi: 10.1016/j.antiviral.2013.03.021. Epub 2013 Apr 9. Antiviral Res. 2013. PMID: 23583287 Free PMC article. Review.
-
Intranasal Inactivated Influenza Vaccines: a Reasonable Approach to Improve the Efficacy of Influenza Vaccine?Jpn J Infect Dis. 2016;69(3):165-79. doi: 10.7883/yoken.JJID.2015.560. Jpn J Infect Dis. 2016. PMID: 27212584 Review.
Cited by
-
A Next-Generation Adenoviral Vaccine Elicits Mucosal and Systemic Immunogenicity and Reduces Viral Shedding after SARS-CoV-2 Challenge in Nonhuman Primates.Vaccines (Basel). 2024 Jan 27;12(2):132. doi: 10.3390/vaccines12020132. Vaccines (Basel). 2024. PMID: 38400116 Free PMC article.
-
Association between selective IgA deficiency and COVID-19.J Clin Biochem Nutr. 2020 Sep;67(2):122-125. doi: 10.3164/jcbn.20-102. Epub 2020 Aug 21. J Clin Biochem Nutr. 2020. PMID: 33041507 Free PMC article.
-
The structure of the teleost Immunoglobulin M core provides insights on polymeric antibody evolution, assembly, and function.Nat Commun. 2023 Nov 21;14(1):7583. doi: 10.1038/s41467-023-43240-z. Nat Commun. 2023. PMID: 37989996 Free PMC article.
-
How to assess the effectiveness of nasal influenza vaccines? Role and measurement of sIgA in mucosal secretions.Influenza Other Respir Viruses. 2019 Sep;13(5):429-437. doi: 10.1111/irv.12664. Epub 2019 Jun 21. Influenza Other Respir Viruses. 2019. PMID: 31225704 Free PMC article. Review.
-
Experimental Efficacy of an Alphavirus Vectored RNA Particle Vaccine Against Porcine Parainfluenza Virus-1 in Conventional Weaned Pigs.Viruses. 2025 Apr 14;17(4):565. doi: 10.3390/v17040565. Viruses. 2025. PMID: 40285006 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
