

The Multiple Biological Targets of Hops and Bioactive Compounds
- PMID: 30608650
- PMCID: PMC6643004
- DOI: 10.1021/acs.chemrestox.8b00345
The Multiple Biological Targets of Hops and Bioactive Compounds
Erratum in
-
Correction to The Multiple Biological Targets of Hops and Bioactive Compounds.Chem Res Toxicol. 2019 Aug 19;32(8):1732. doi: 10.1021/acs.chemrestox.9b00300. Epub 2019 Aug 8. Chem Res Toxicol. 2019. PMID: 31392882 No abstract available.
Abstract
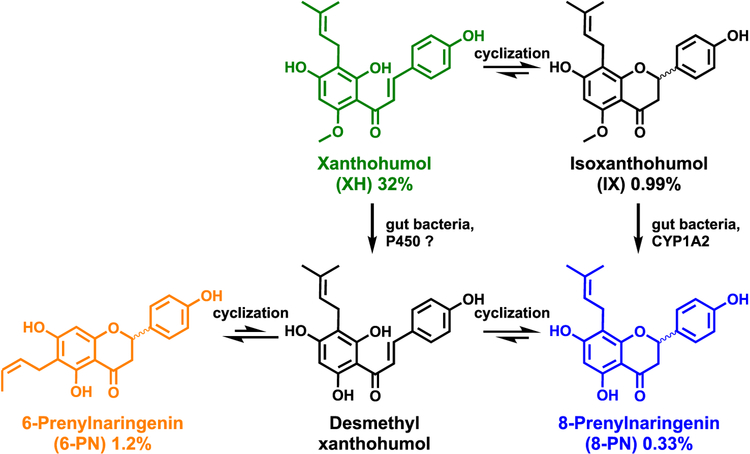
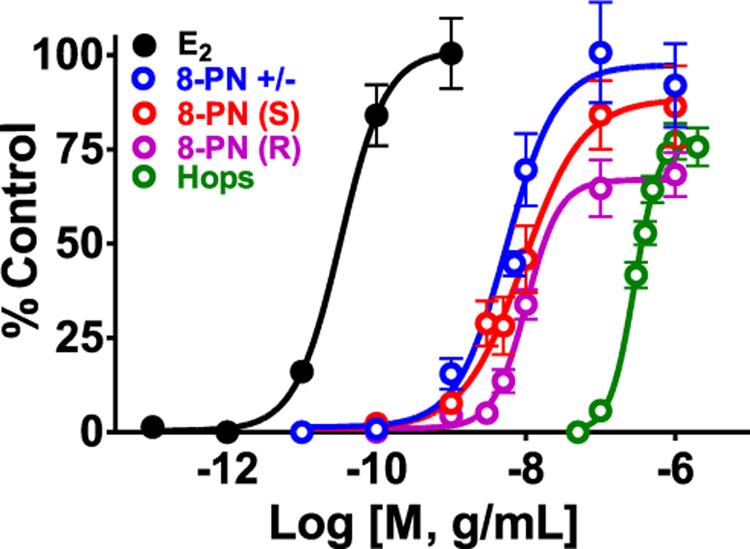
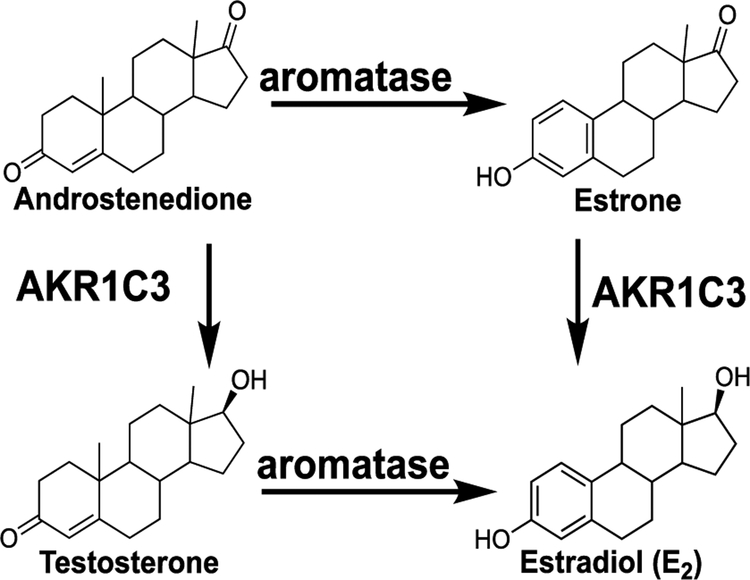
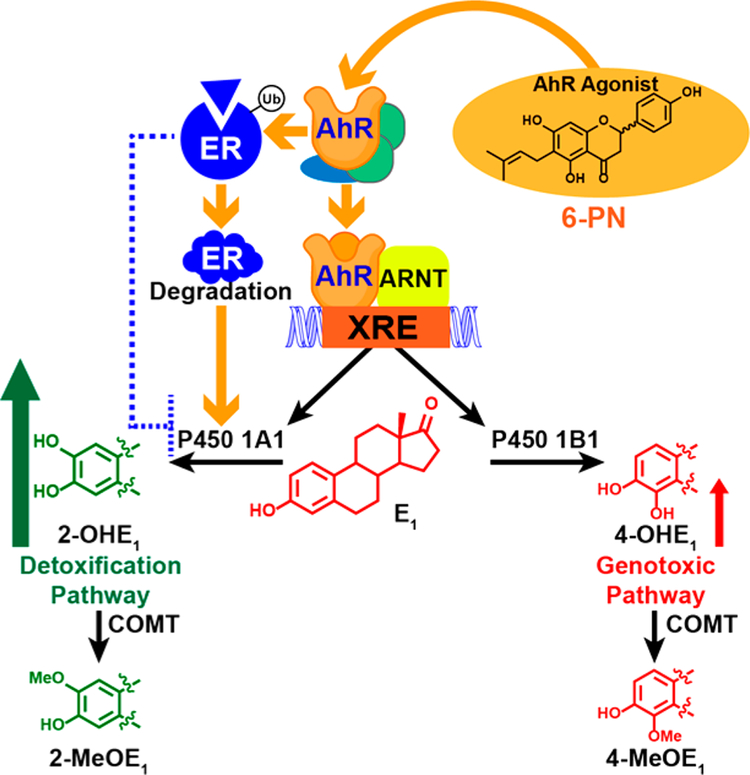
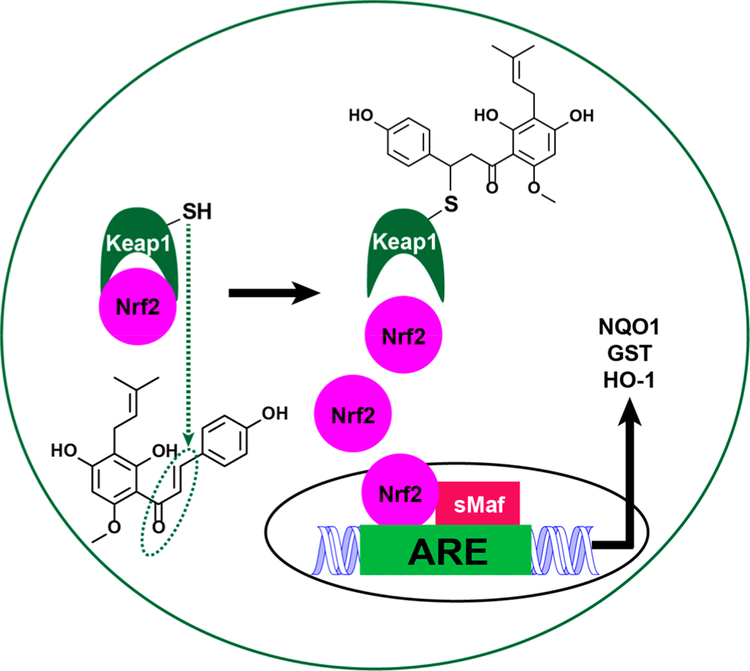
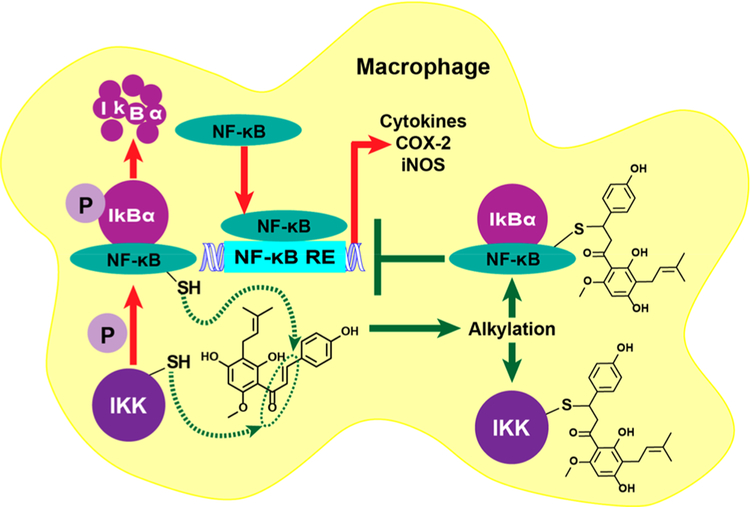
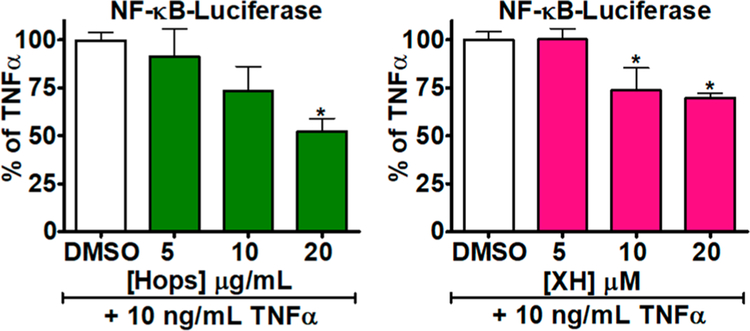
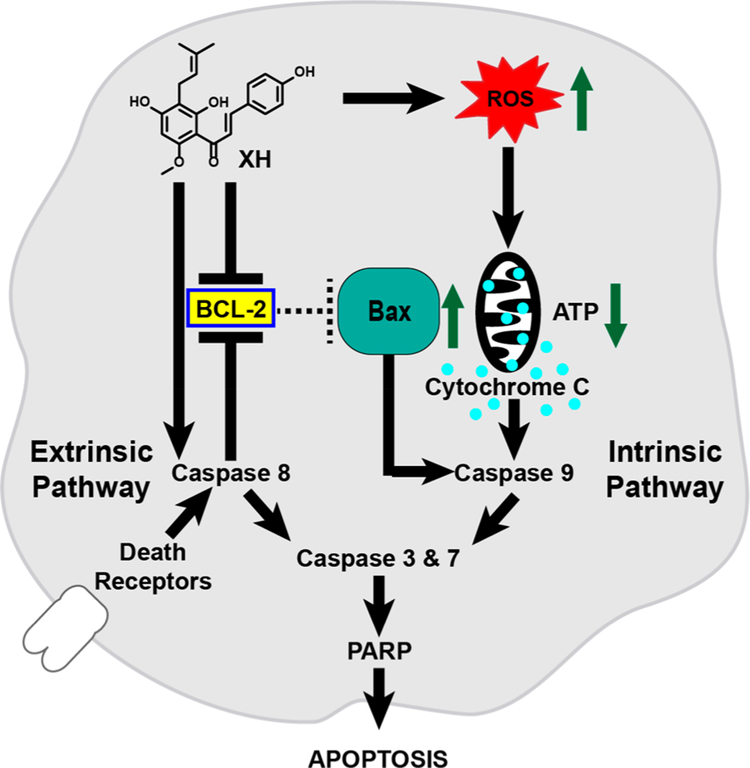
Botanical dietary supplements for women's health are increasingly popular. Older women tend to take botanical supplements such as hops as natural alternatives to traditional hormone therapy to relieve menopausal symptoms. Especially extracts from spent hops, the plant material remaining after beer brewing, are enriched in bioactive prenylated flavonoids that correlate with the health benefits of the plant. The chalcone xanthohumol (XH) is the major prenylated flavonoid in spent hops. Other less abundant but important bioactive prenylated flavonoids are isoxanthohumol (IX), 8-prenylnaringenin (8-PN), and 6-prenylnaringenin (6-PN). Pharmacokinetic studies revealed that these flavonoids are conjugated rapidly with glucuronic acid. XH also undergoes phase I metabolism in vivo to form IX, 8-PN, and 6-PN. Several hop constituents are responsible for distinct effects linked to multiple biological targets, including hormonal, metabolic, inflammatory, and epigenetic pathways. 8-PN is one of the most potent phytoestrogens and is responsible for hops' estrogenic activities. Hops also inhibit aromatase activity, which is linked to 8-PN. The weak electrophile, XH, can activate the Keap1-Nrf2 pathway and turn on the synthesis of detoxification enzymes such as NAD(P)H-quinone oxidoreductase 1 and glutathione S-transferase. XH also alkylates IKK and NF-κB, resulting in anti-inflammatory activity. Antiobesity activities have been described for XH and XH-rich hop extracts likely through activation of AMP-activated protein kinase signaling pathways. Hop extracts modulate the estrogen chemical carcinogenesis pathway by enhancing P450 1A1 detoxification. The mechanism appears to involve activation of the aryl hydrocarbon receptor (AhR) by the AhR agonist, 6-PN, leading to degradation of the estrogen receptor. Finally, prenylated phenols from hops are known inhibitors of P450 1A1/2; P450 1B1; and P450 2C8, 2C9, and 2C19. Understanding the biological targets of hop dietary supplements and their phytoconstituents will ultimately lead to standardized botanical products with higher efficacy, safety, and chemopreventive properties.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





References
-
- Stevens JF, and Page JE (2004) Xanthohumol and related prenylflavonoids from hops and beer: to your good health! Phytochemistry 65, 1317–1330. - PubMed
-
- Karabin M, Hudcova T, Jelinek L, and Dostalek P (2016) Biologically active compounds from hops and prospects for their use. Compr. Rev. Food Sci. Food Saf 15, 542–567. - PubMed
-
- Karabin M, Hudcova T, Jelinek L, and Dostalek P (2015) Biotransformations and biological activities of hop flavonoids. Biotechnol. Adv. 33, 1063–1090. - PubMed
-
- Aghamiri V, Mirghafourvand M, Mohammad-Alizadeh-Charandabi S, and Nazemiyeh H (2016) The effect of Hop (Humulus lupulus L.) on early menopausal symptoms and hot flashes: A randomized placebo-controlled trial. Complement. Ther. Clin. Pract 23, 130–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
