

Thymic expression of IL-4 and IL-15 after systemic inflammatory or infectious Th1 disease processes induce the acquisition of "innate" characteristics during CD8+ T cell development
- PMID: 30608984
- PMCID: PMC6319713
- DOI: 10.1371/journal.ppat.1007456
Thymic expression of IL-4 and IL-15 after systemic inflammatory or infectious Th1 disease processes induce the acquisition of "innate" characteristics during CD8+ T cell development
Abstract
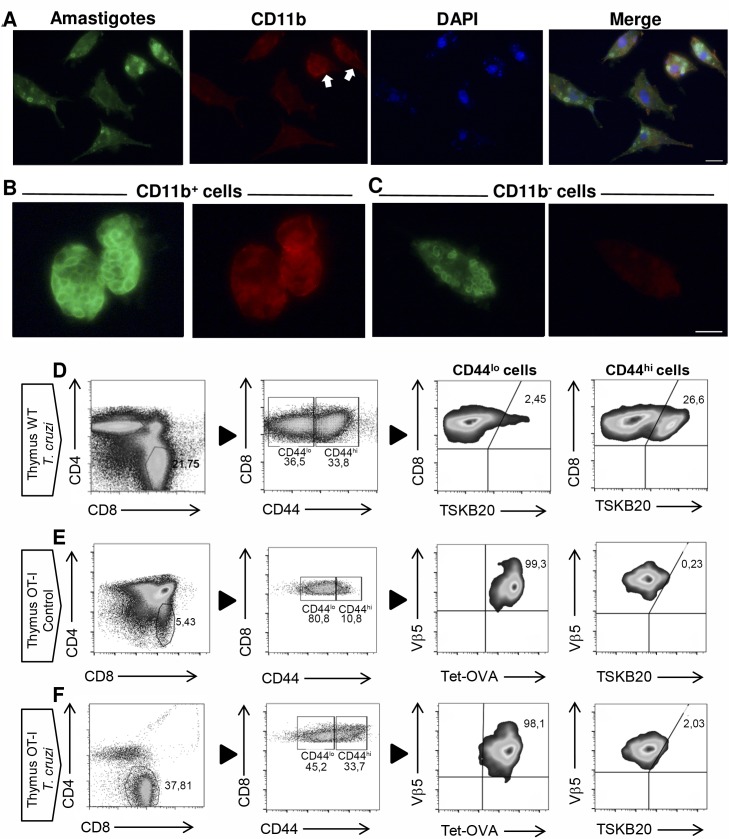
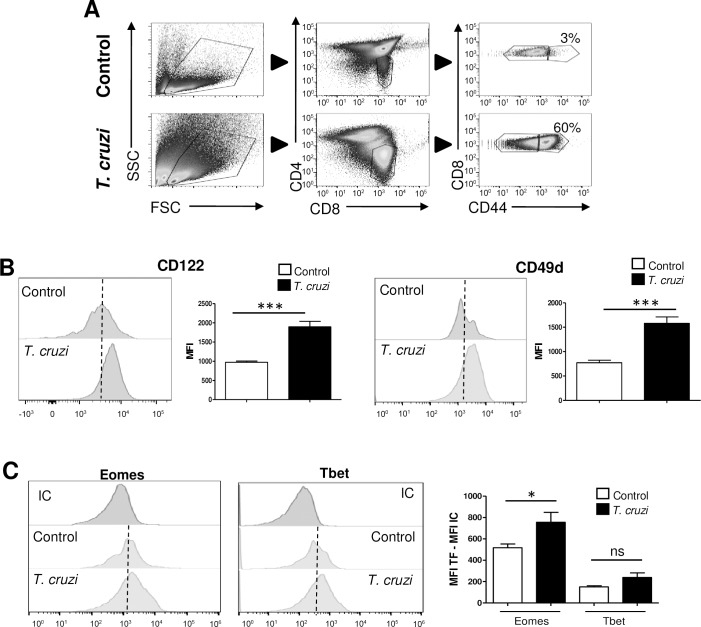
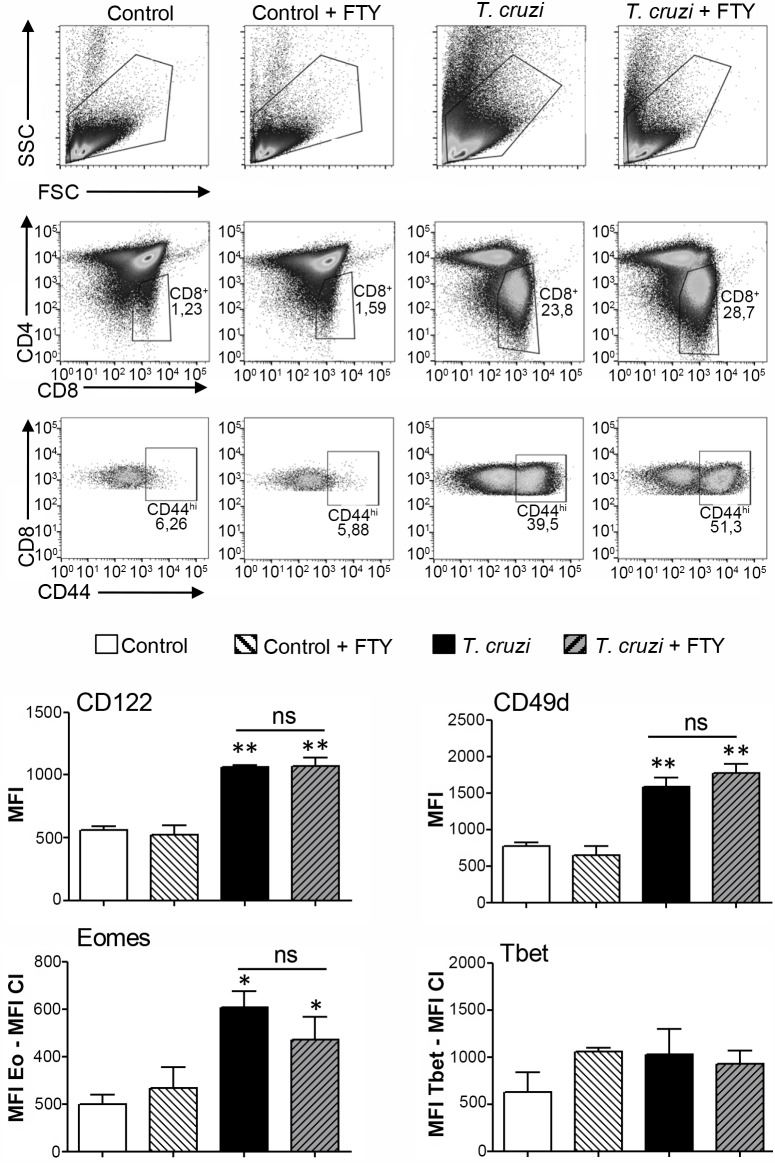
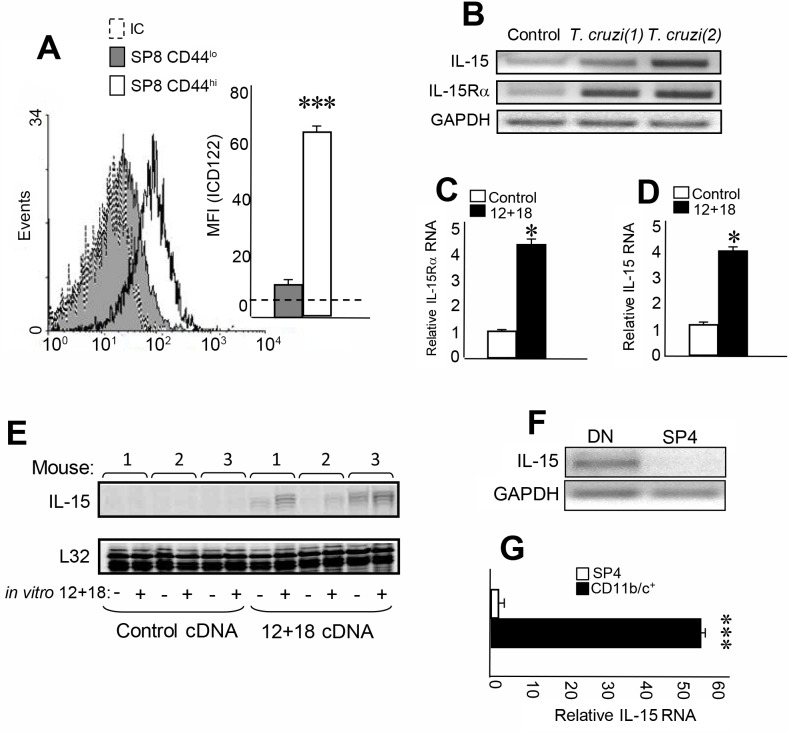
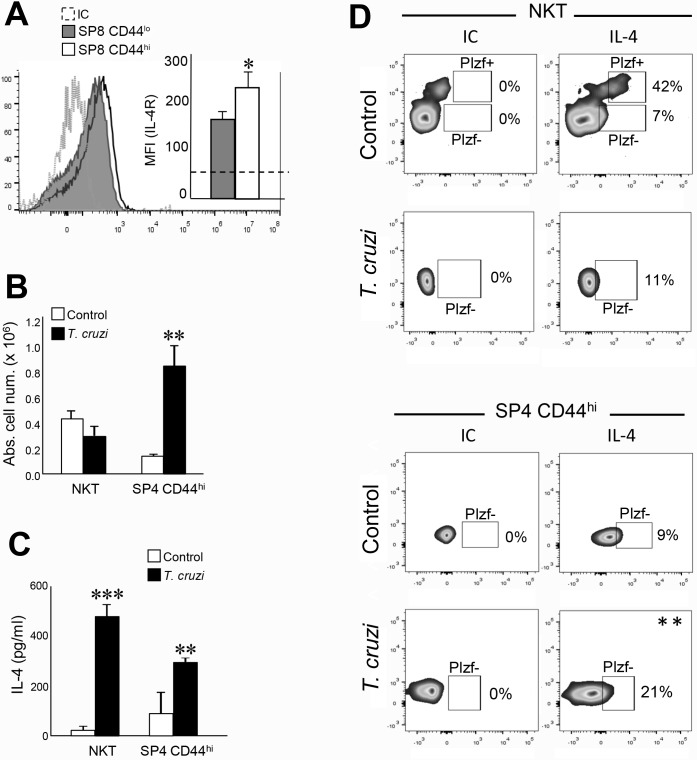
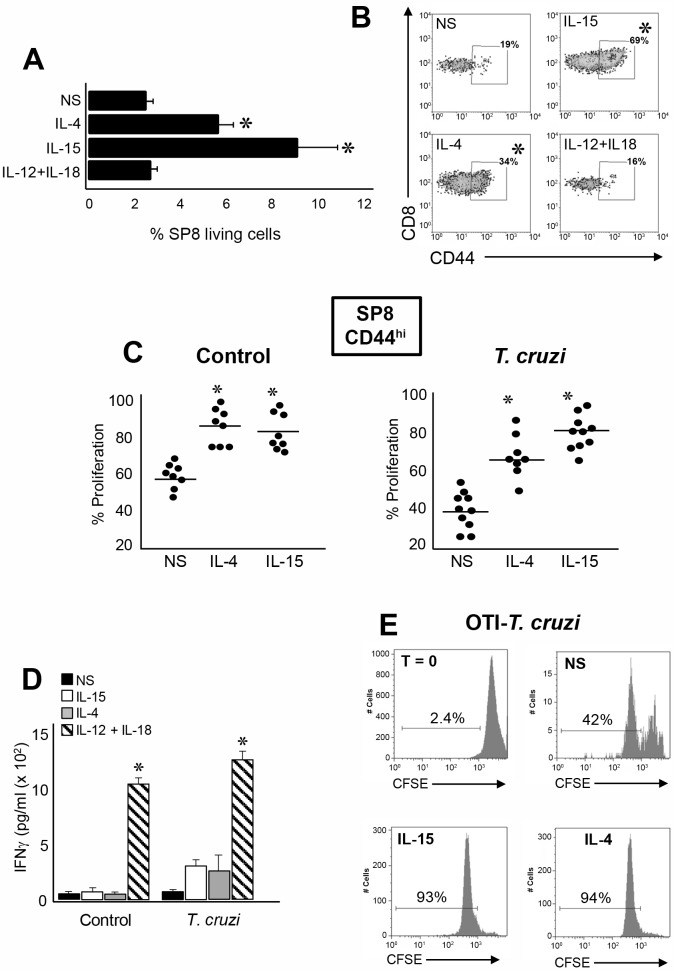
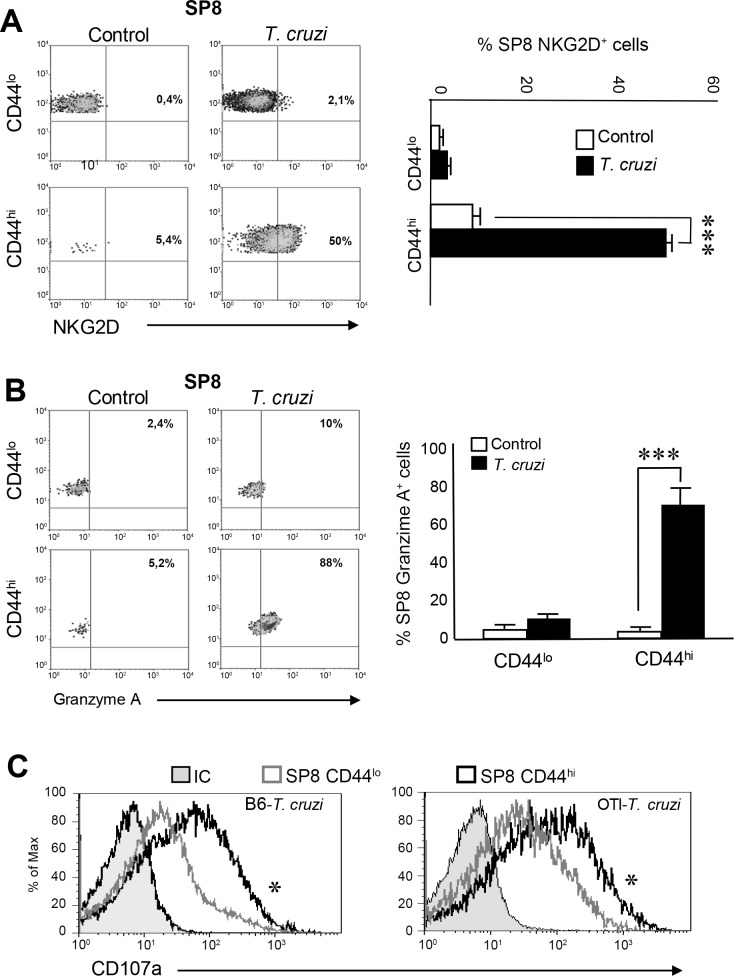
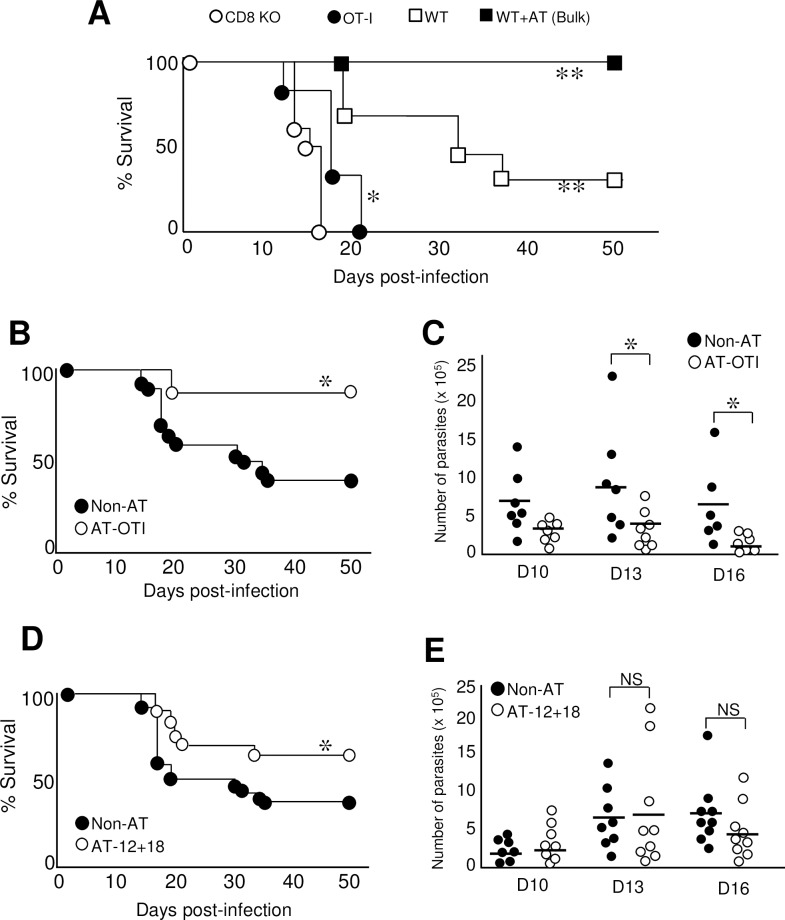
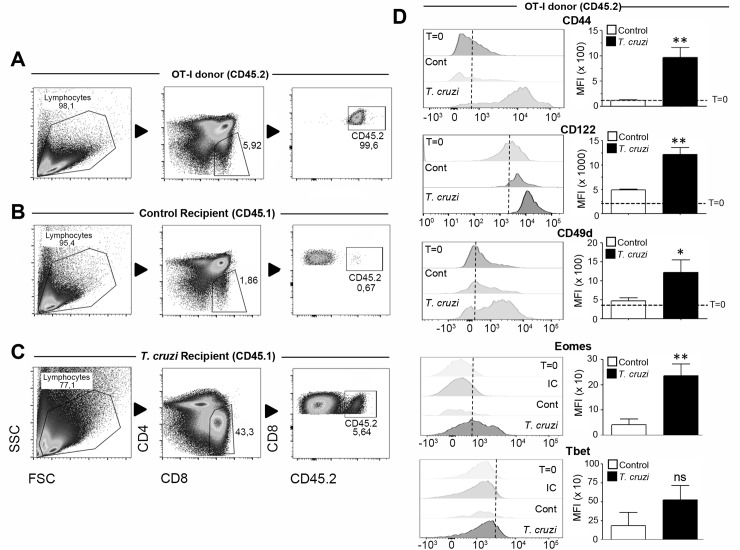
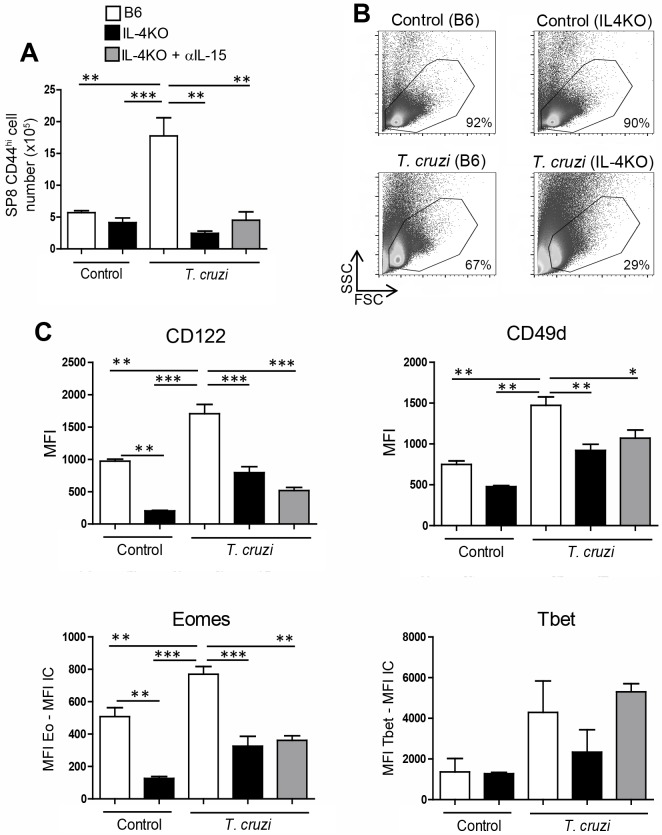
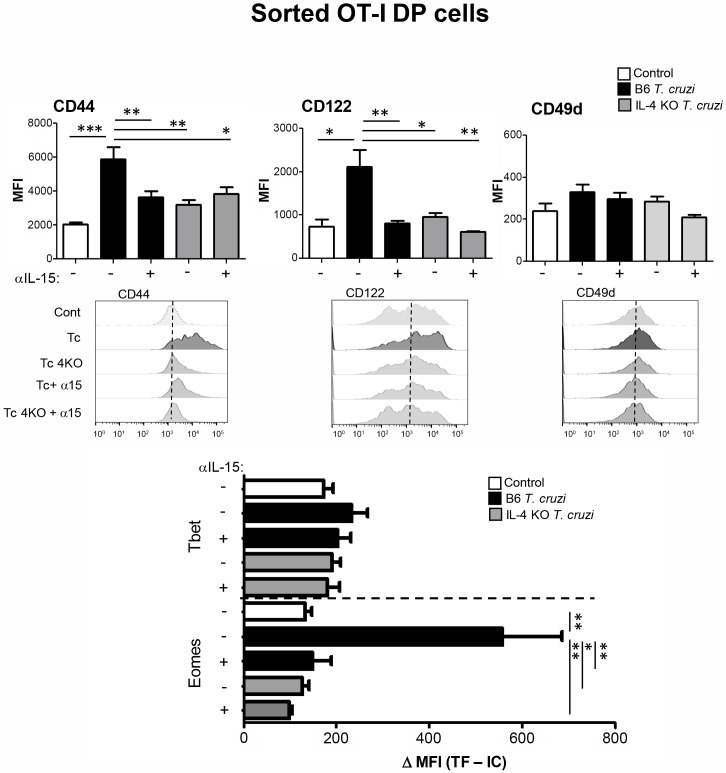
Innate CD8+ T cells express a memory-like phenotype and demonstrate a strong cytotoxic capacity that is critical during the early phase of the host response to certain bacterial and viral infections. These cells arise in the thymus and depend on IL-4 and IL-15 for their development. Even though innate CD8+ T cells exist in the thymus of WT mice in low numbers, they are highly enriched in KO mice that lack certain kinases, leading to an increase in IL-4 production by thymic NKT cells. Our work describes that in C57BL/6 WT mice undergoing a Th1 biased infectious disease, the thymus experiences an enrichment of single positive CD8 (SP8) thymocytes that share all the established phenotypical and functional characteristics of innate CD8+ T cells. Moreover, through in vivo experiments, we demonstrate a significant increase in survival and a lower parasitemia in mice adoptively transferred with SP8 thymocytes from OT I-T. cruzi-infected mice, demonstrating that innate CD8+ thymocytes are able to protect against a lethal T. cruzi infection in an Ag-independent manner. Interestingly, we obtained similar results when using thymocytes from systemic IL-12 + IL-18-treated mice. This data indicates that cytokines triggered during the acute stage of a Th1 infectious process induce thymic production of IL-4 along with IL-15 expression resulting in an adequate niche for development of innate CD8+ T cells as early as the double positive (DP) stage. Our data demonstrate that the thymus can sense systemic inflammatory situations and alter its conventional CD8 developmental pathway when a rapid innate immune response is required to control different types of pathogens.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
