

Neuronal sub-compartmentalization: a strategy to optimize neuronal function
- PMID: 30609235
- PMCID: PMC6617802
- DOI: 10.1111/brv.12487
Neuronal sub-compartmentalization: a strategy to optimize neuronal function
Abstract
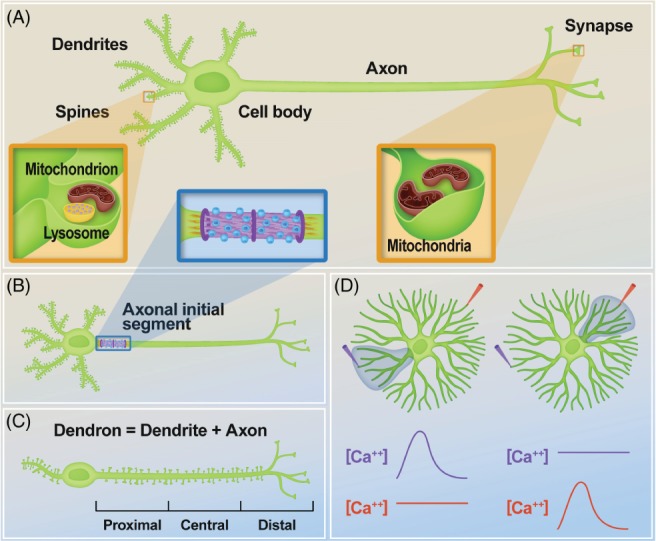
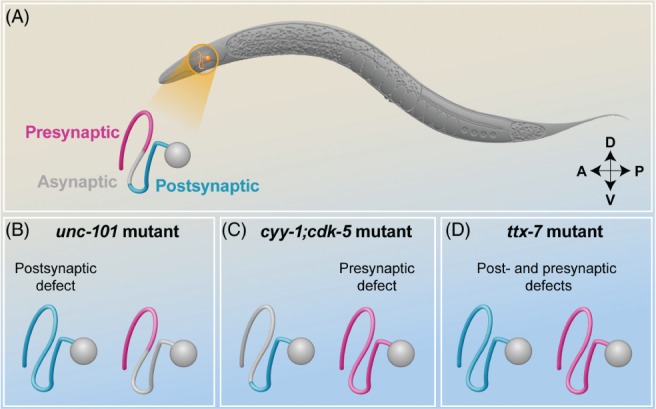
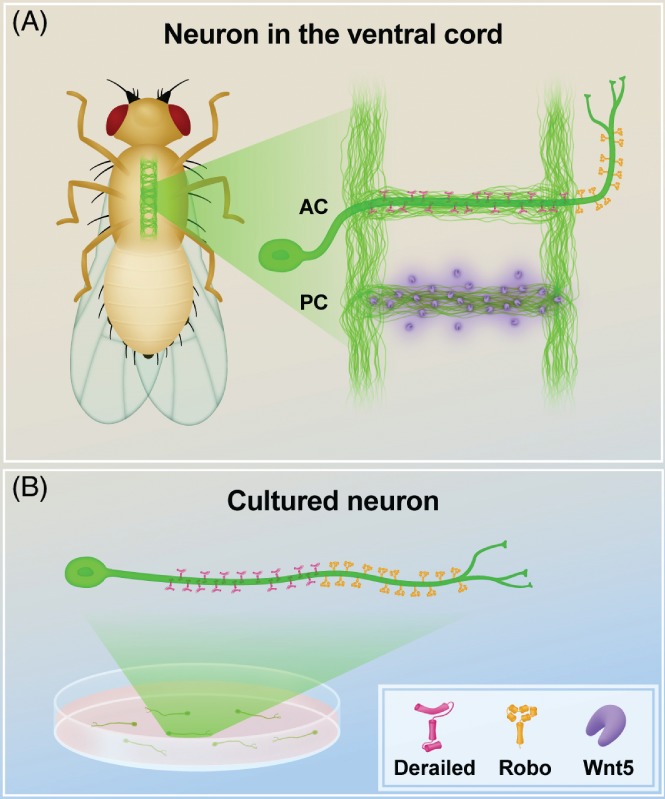
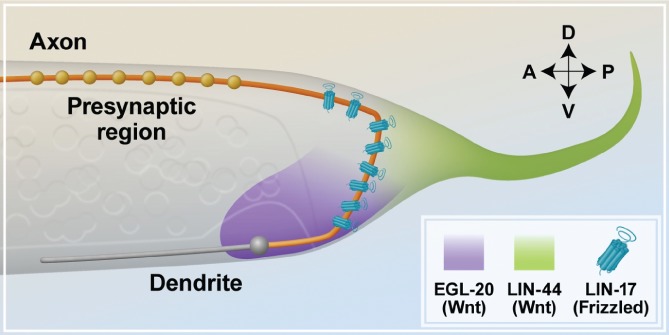
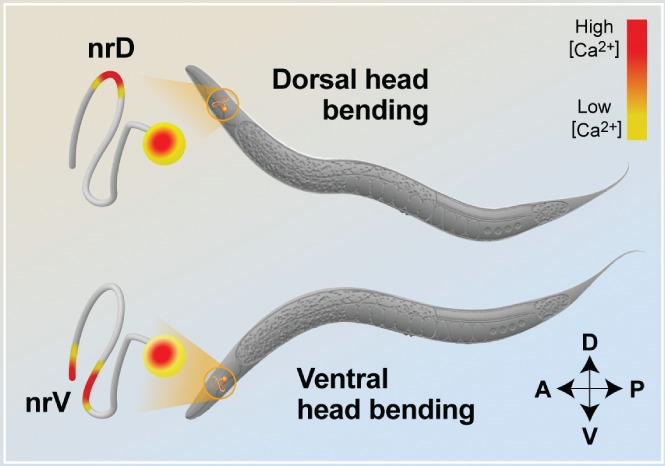
Neurons are highly polarized cells that consist of three main structural and functional domains: a cell body or soma, an axon, and dendrites. These domains contain smaller compartments with essential roles for proper neuronal function, such as the axonal presynaptic boutons and the dendritic postsynaptic spines. The structure and function of these compartments have now been characterized in great detail. Intriguingly, however, in the last decade additional levels of compartmentalization within the axon and the dendrites have been identified, revealing that these structures are much more complex than previously thought. Herein we examine several types of structural and functional sub-compartmentalization found in neurons of both vertebrates and invertebrates. For example, in mammalian neurons the axonal initial segment functions as a sub-compartment to initiate the action potential, to select molecules passing into the axon, and to maintain neuronal polarization. Moreover, work in Drosophila melanogaster has shown that two distinct axonal guidance receptors are precisely clustered in adjacent segments of the commissural axons both in vivo and in vitro, suggesting a cell-intrinsic mechanism underlying the compartmentalized receptor localization. In Caenorhabditis elegans, a subset of interneurons exhibits calcium dynamics that are localized to specific sections of the axon and control the gait of navigation, demonstrating a regulatory role of compartmentalized neuronal activity in behaviour. These findings have led to a number of new questions, which are important for our understanding of neuronal development and function. How are these sub-compartments established and maintained? What molecular machinery and cellular events are involved? What is their functional significance for the neuron? Here, we reflect on these and other key questions that remain to be addressed in this expanding field of biology.
Keywords: axonal and dendritic sub-compartments; compartmentalization; neuronal activity; neuronal development; patterning; polarization; unipolar neurons.
© 2019 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures
Similar articles
-
Polarity and intracellular compartmentalization of Drosophila neurons.Neural Dev. 2007 Apr 30;2:7. doi: 10.1186/1749-8104-2-7. Neural Dev. 2007. PMID: 17470283 Free PMC article.
-
Compartmentalization within neurites: its mechanisms and implications.Dev Neurobiol. 2011 Jun;71(6):458-73. doi: 10.1002/dneu.20859. Dev Neurobiol. 2011. PMID: 21557500 Review.
-
Axon outgrowth and neuronal differentiation defects after a-SMN and FL-SMN silencing in primary hippocampal cultures.PLoS One. 2018 Jun 14;13(6):e0199105. doi: 10.1371/journal.pone.0199105. eCollection 2018. PLoS One. 2018. PMID: 29902268 Free PMC article.
-
Intra-axonal patterning: intrinsic compartmentalization of the axonal membrane in Drosophila neurons.Neuron. 2009 Oct 29;64(2):188-99. doi: 10.1016/j.neuron.2009.08.019. Neuron. 2009. PMID: 19874787
-
Neuronal polarity and trafficking.Neuron. 2003 Oct 9;40(2):277-95. doi: 10.1016/s0896-6273(03)00629-9. Neuron. 2003. PMID: 14556709 Review.
Cited by
-
Intrinsic and extrinsic mechanisms of synapse formation and specificity in C. elegans.Cell Mol Life Sci. 2019 Jul;76(14):2719-2738. doi: 10.1007/s00018-019-03109-1. Epub 2019 Apr 29. Cell Mol Life Sci. 2019. PMID: 31037336 Free PMC article. Review.
-
Reactive Oxygen Species Modulate Activity-Dependent AMPA Receptor Transport in C. elegans.J Neurosci. 2020 Sep 23;40(39):7405-7420. doi: 10.1523/JNEUROSCI.0902-20.2020. Epub 2020 Aug 26. J Neurosci. 2020. PMID: 32847966 Free PMC article.
-
Neuromedin U signaling regulates retrieval of learned salt avoidance in a C. elegans gustatory circuit.Nat Commun. 2020 Apr 29;11(1):2076. doi: 10.1038/s41467-020-15964-9. Nat Commun. 2020. PMID: 32350283 Free PMC article.
-
An integrative data-driven model simulating C. elegans brain, body and environment interactions.Nat Comput Sci. 2024 Dec;4(12):978-990. doi: 10.1038/s43588-024-00738-w. Epub 2024 Dec 16. Nat Comput Sci. 2024. PMID: 39681671 Free PMC article.
-
The synaptic organization in the Caenorhabditis elegans neural network suggests significant local compartmentalized computations.Proc Natl Acad Sci U S A. 2023 Jan 17;120(3):e2201699120. doi: 10.1073/pnas.2201699120. Epub 2023 Jan 11. Proc Natl Acad Sci U S A. 2023. PMID: 36630454 Free PMC article.
References
-
- Antar, L. N. , Afroz, R. , Dictenberg, J. B. , Carroll, R. C. & Bassell, G. J. (2004). Metabotropic glutamate receptor activation regulates Fragile X mental retardation protein and Fmr1 mRNA localization differentially in dendrites and at synapses. The Journal of Neuroscience 24, 2648–2655. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
