

Evolution, phylogenetic distribution and functional ecology of division of labour in trematodes
- PMID: 30609937
- PMCID: PMC6320615
- DOI: 10.1186/s13071-018-3241-6
Evolution, phylogenetic distribution and functional ecology of division of labour in trematodes
Abstract
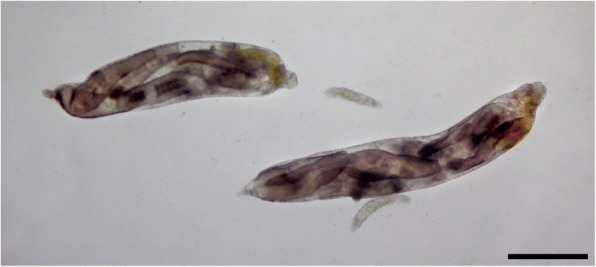
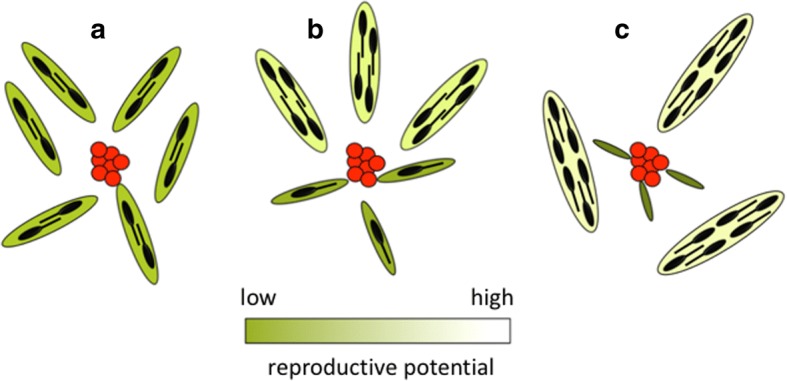
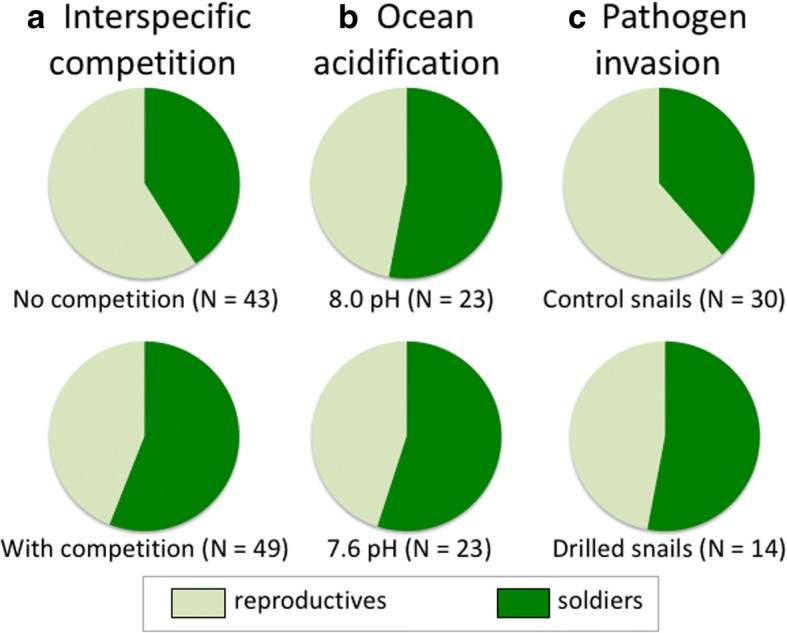
Division of labour has evolved in many social animals where colonies consist of clones or close kin. It involves the performance of different tasks by morphologically distinct castes, leading to increased colony fitness. Recently, a form of division of labour has been discovered in trematodes: clonal rediae inside the snail intermediate host belong either to a large-bodied reproductive caste, or to a much smaller and morphologically distinct 'soldier' caste which defends the colony against co-infecting trematodes. We review recent research on this phenomenon, focusing on its phylogenetic distribution, its possible evolutionary origins, and how division of labour functions to allow trematode colonies within their snail host to adjust to threats and changing conditions. To date, division of labour has been documented in 15 species from three families: Himasthlidae, Philophthalmidae and Heterophyidae. Although this list of species is certainly incomplete, the evidence suggests that division of labour has arisen independently more than once in the evolutionary history of trematodes. We propose a simple scenario for the gradual evolution of division of labour in trematodes facing a high risk of competition in a long-lived snail host. Starting with initial conditions prior to the origin of castes (size variation among rediae within a colony, size-dependent production of cercariae by rediae, and a trade-off between cercarial production and other functions, such as defence), maximising colony fitness (R0) can lead to caste formation or the age-structured division of labour observed in some trematodes. Finally, we summarise recent research showing that caste ratios, i.e. relative numbers of reproductive and soldier rediae per colony, become more soldier-biased in colonies exposed to competition from another trematode species sharing the same snail, and also respond to other stressors threatening the host's survival or the colony itself. In addition, there is evidence of asymmetrical phenotypic plasticity among individual caste members: reproductives can assume defensive functions against competitors in the absence of soldiers, whereas soldiers are incapable of growing into reproductives if the latter's numbers are reduced. We conclude by highlighting future research directions, and the advantages of trematodes as model systems to study social evolution.
Keywords: Caste; Cercariae; Fitness; Multiple infections; Philophthalmus; Rediae; Sociality; Soldiers; Trade-off.
Conflict of interest statement
Ethics approval
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Caste ratio adjustments in response to perceived and realised competition in parasites with division of labour.J Anim Ecol. 2018 Sep;87(5):1429-1439. doi: 10.1111/1365-2656.12873. Epub 2018 Jun 29. J Anim Ecol. 2018. PMID: 29888446
-
Trematodes with a reproductive division of labour: heterophyids also have a soldier caste and early infections reveal how colonies become structured.Int J Parasitol. 2017 Jan;47(1):41-50. doi: 10.1016/j.ijpara.2016.10.003. Epub 2016 Nov 30. Int J Parasitol. 2017. PMID: 27914977
-
Reproduction and caste ratios under stress in trematode colonies with a division of labour.Parasitology. 2013 Jun;140(7):825-32. doi: 10.1017/S0031182012002235. Epub 2013 Feb 27. Parasitology. 2013. PMID: 23442525
-
Ecology of larval trematodes in three marine gastropods.Parasitology. 2002;124 Suppl:S43-56. doi: 10.1017/s0031182002001452. Parasitology. 2002. PMID: 12396215 Review.
-
Chronobiology of trematode cercarial emergence: from data recovery to epidemiological, ecological and evolutionary implications.Adv Parasitol. 2015 Apr;88:123-64. doi: 10.1016/bs.apar.2015.02.003. Epub 2015 Mar 23. Adv Parasitol. 2015. PMID: 25911367 Review.
Cited by
-
When being flexible matters: Ecological underpinnings for the evolution of collective flexibility and task allocation.Proc Natl Acad Sci U S A. 2022 May 3;119(18):e2116066119. doi: 10.1073/pnas.2116066119. Epub 2022 Apr 29. Proc Natl Acad Sci U S A. 2022. PMID: 35486699 Free PMC article.
-
The physical soldier caste of an invasive, human-infecting flatworm is morphologically extreme and obligately sterile.Proc Natl Acad Sci U S A. 2024 Jul 30;121(31):e2400953121. doi: 10.1073/pnas.2400953121. Epub 2024 Jul 23. Proc Natl Acad Sci U S A. 2024. PMID: 39042696 Free PMC article.
-
Social evolution and reproductive castes in trematode parasites.Proc Natl Acad Sci U S A. 2024 Sep 10;121(37):e2414228121. doi: 10.1073/pnas.2414228121. Epub 2024 Sep 3. Proc Natl Acad Sci U S A. 2024. PMID: 39226370 Free PMC article. No abstract available.
-
Freshwater trematodes differ from marine trematodes in patterns connected with division of labor.PeerJ. 2024 Apr 12;12:e17211. doi: 10.7717/peerj.17211. eCollection 2024. PeerJ. 2024. PMID: 38623495 Free PMC article.
-
The weird eusociality of polyembryonic parasites.Biol Lett. 2021 Mar;17(4):20210026. doi: 10.1098/rsbl.2021.0026. Epub 2021 Apr 7. Biol Lett. 2021. PMID: 33823644 Free PMC article.
References
-
- Duarte A, Weissing FJ, Pen I, Keller L. An evolutionary perspective on self-organized division of labor in social insects. Annu Rev Ecol Evol Syst. 2011;42:91–110. doi: 10.1146/annurev-ecolsys-102710-145017. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
