

Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis
- PMID: 30612898
- PMCID: PMC6509403
- DOI: 10.1016/j.cmet.2018.12.008
Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis
Erratum in
-
Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis.Cell Metab. 2019 May 7;29(5):1233. doi: 10.1016/j.cmet.2019.01.013. Cell Metab. 2019. PMID: 31067450 Free PMC article. No abstract available.
Abstract
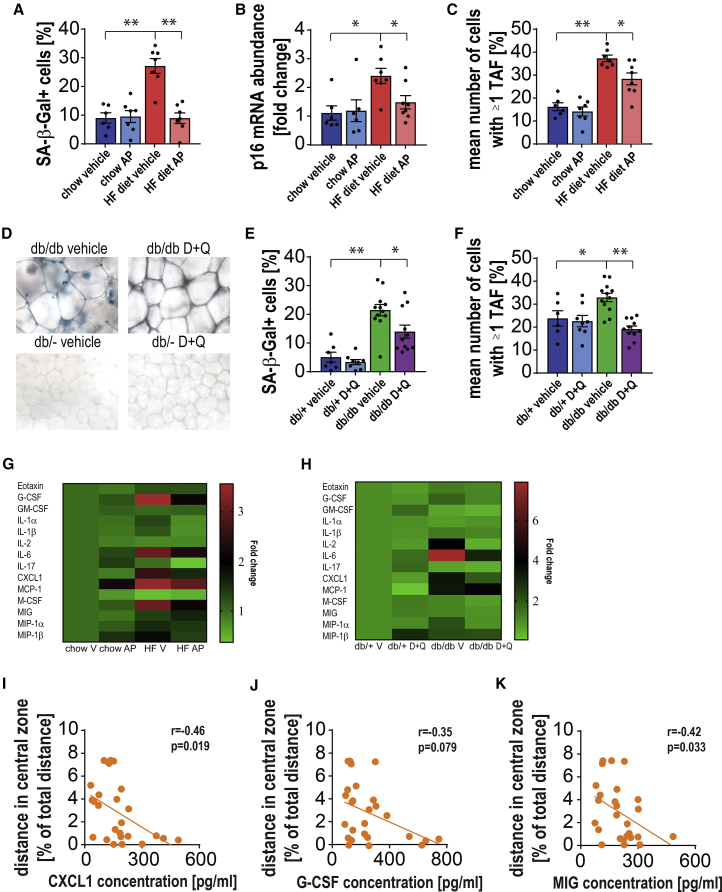
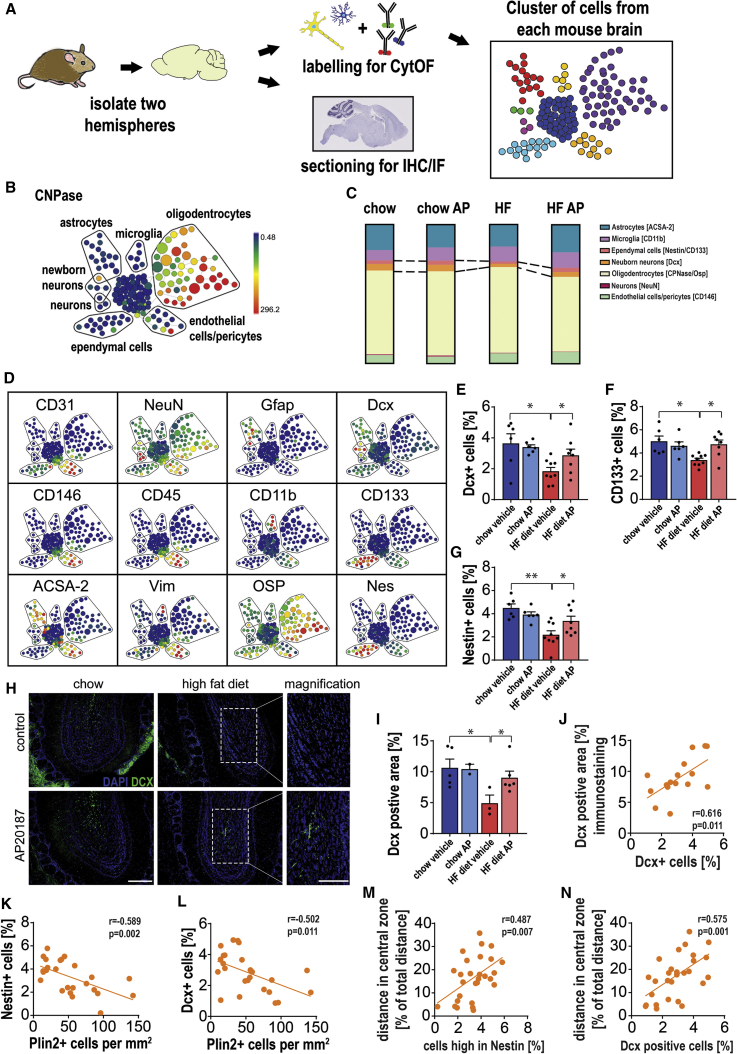
Cellular senescence entails a stable cell-cycle arrest and a pro-inflammatory secretory phenotype, which contributes to aging and age-related diseases. Obesity is associated with increased senescent cell burden and neuropsychiatric disorders, including anxiety and depression. To investigate the role of senescence in obesity-related neuropsychiatric dysfunction, we used the INK-ATTAC mouse model, from which p16Ink4a-expressing senescent cells can be eliminated, and senolytic drugs dasatinib and quercetin. We found that obesity results in the accumulation of senescent glial cells in proximity to the lateral ventricle, a region in which adult neurogenesis occurs. Furthermore, senescent glial cells exhibit excessive fat deposits, a phenotype we termed "accumulation of lipids in senescence." Clearing senescent cells from high fat-fed or leptin receptor-deficient obese mice restored neurogenesis and alleviated anxiety-related behavior. Our study provides proof-of-concept evidence that senescent cells are major contributors to obesity-induced anxiety and that senolytics are a potential new therapeutic avenue for treating neuropsychiatric disorders.
Keywords: aging; anxiety; anxiety-like behavior; brain; high-fat diet; lipid droplets; neurogenesis; obesity; senescence; stem cells.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
- BB/H022384/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R37 AG013925/AG/NIA NIH HHS/United States
- P30 AG013319/AG/NIA NIH HHS/United States
- BB/F010966/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 24009/CRUK_/Cancer Research UK/United Kingdom
- BB/S006710/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/I020748/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/P020941/1/MRC_/Medical Research Council/United Kingdom
- MR/L016354/1/MRC_/Medical Research Council/United Kingdom
- R01 AG013925/AG/NIA NIH HHS/United States
- C12161/A24009/CRUK_/Cancer Research UK/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
