

Hyper-Editing of Cell-Cycle Regulatory and Tumor Suppressor RNA Promotes Malignant Progenitor Propagation
- PMID: 30612940
- PMCID: PMC6333511
- DOI: 10.1016/j.ccell.2018.11.017
Hyper-Editing of Cell-Cycle Regulatory and Tumor Suppressor RNA Promotes Malignant Progenitor Propagation
Abstract
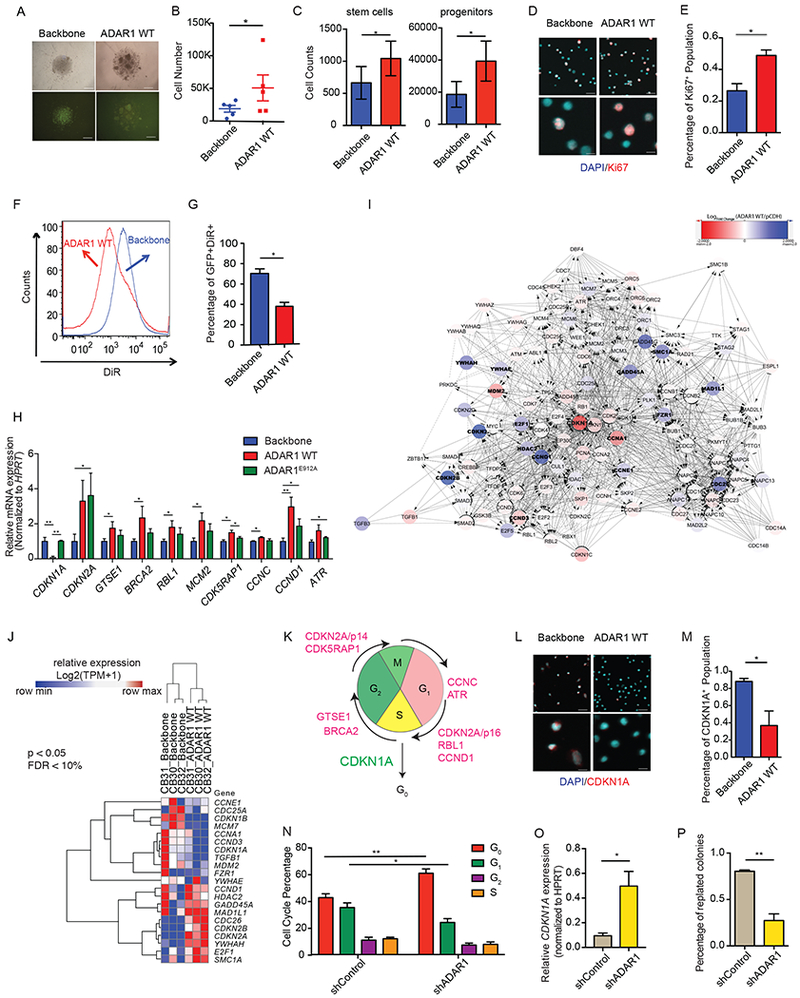
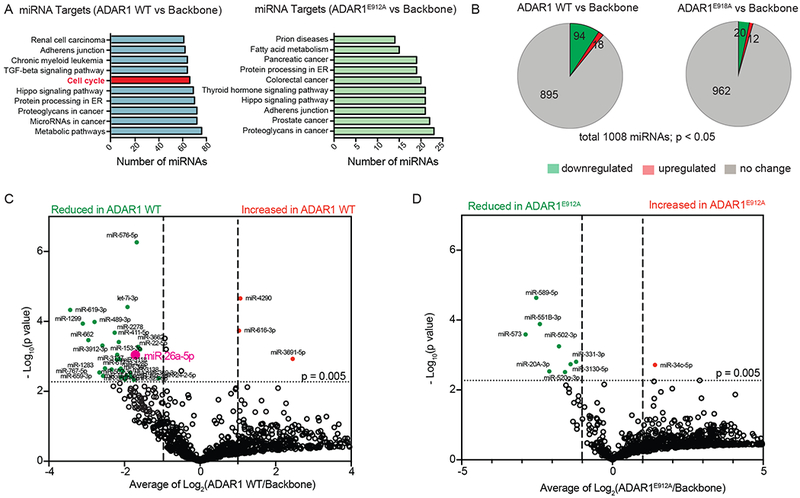
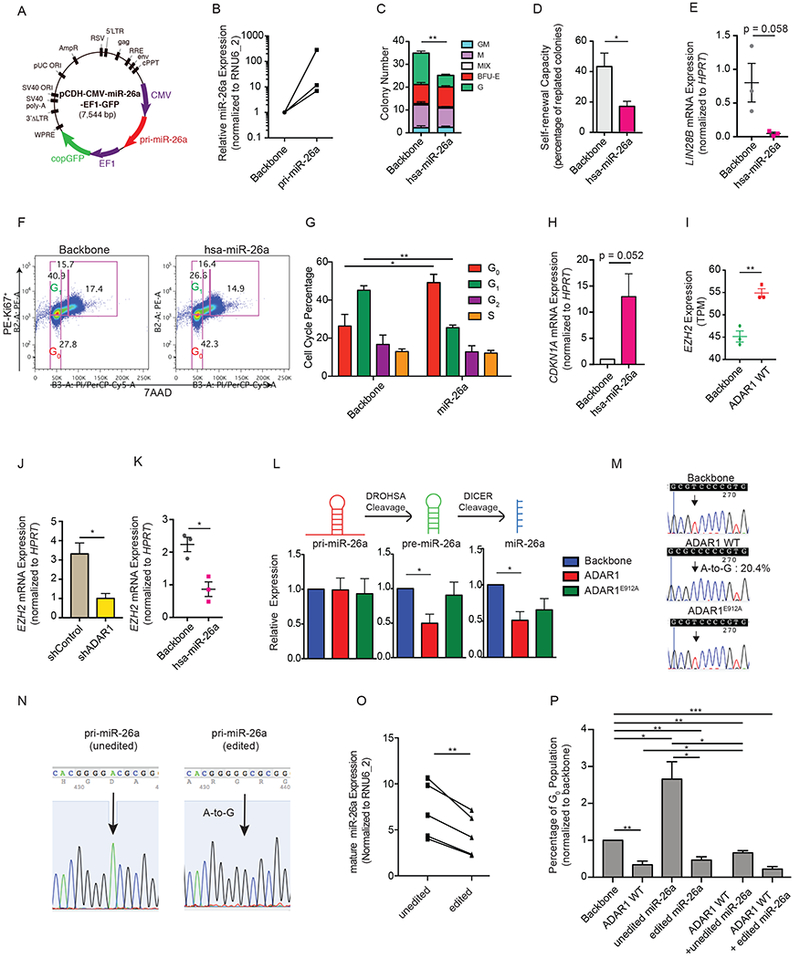
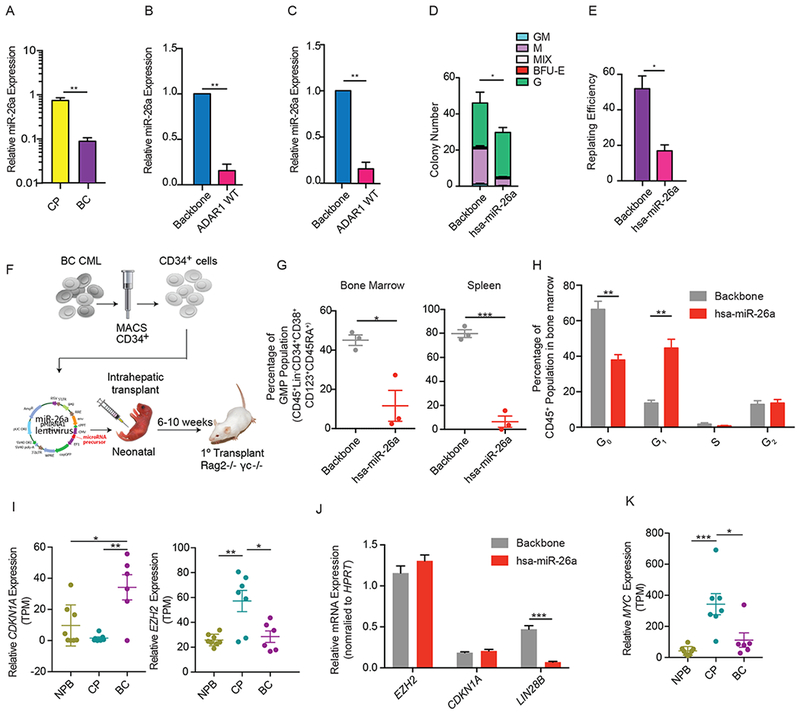
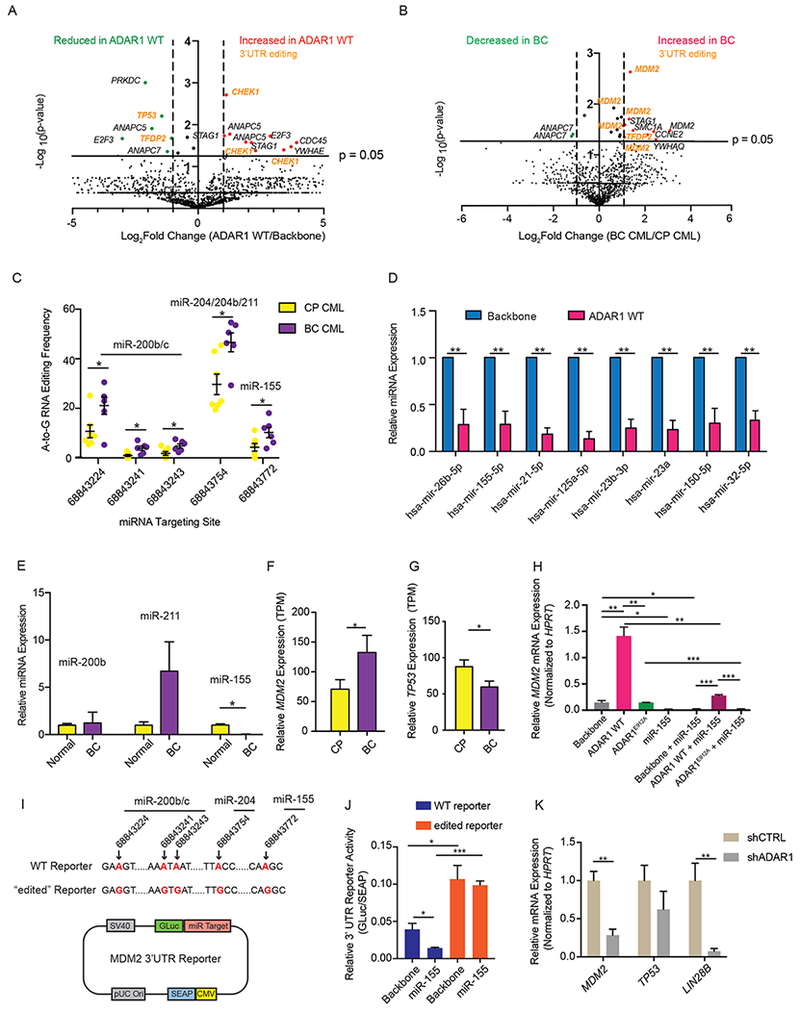
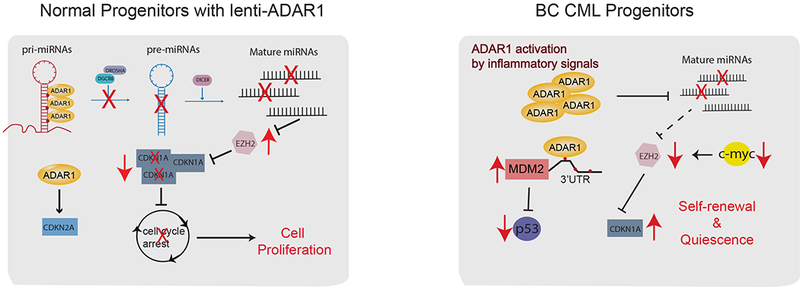
Adenosine deaminase associated with RNA1 (ADAR1) deregulation contributes to therapeutic resistance in many malignancies. Here we show that ADAR1-induced hyper-editing in normal human hematopoietic progenitors impairs miR-26a maturation, which represses CDKN1A expression indirectly via EZH2, thereby accelerating cell-cycle transit. However, in blast crisis chronic myeloid leukemia progenitors, loss of EZH2 expression and increased CDKN1A oppose cell-cycle transit. Moreover, A-to-I editing of both the MDM2 regulatory microRNA and its binding site within the 3' UTR region stabilizes MDM2 transcripts, thereby enhancing blast crisis progenitor propagation. These data reveal a dual mechanism governing malignant transformation of progenitors that is predicated on hyper-editing of cell-cycle-regulatory miRNAs and the 3' UTR binding site of tumor suppressor miRNAs.
Keywords: 3′ UTR; ADAR1; RNA hyper-editing; cancer stem cell; cell cycle; chronic myeloid leukemia; epitranscriptome; leukemia; microRNAs; progenitors; self-renewal.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
C. Jamieson was a co-founder of Impact Biomedicines Inc, which was acquired by Celgene, and holds stock in Forty Seven Inc.
Figures
References
-
- Abrahamsson AE, Geron I, Gotlib J, Dao KH, Barroga CF, Newton IG, Giles FJ, Durocher J, Creusot RS, Karimi M, et al. (2009). Glycogen synthase kinase 3beta missplicing contributes to leukemia stem cell generation. Proceedings of the National Academy of Sciences of the United States of America 106, 3925–3929. - PMC - PubMed
-
- Batzer MA, and Deininger PL (2002). Alu repeats and human genomic diversity. Nat Rev Genet 3, 370–379. - PubMed
-
- Beguelin W, Teater M, Gearhart MD, Calvo Fernandez MT, Goldstein RL, Cardenas MG, Hatzi K, Rosen M, Shen H, Corcoran CM, et al. (2016). EZH2 and BCL6 Cooperate to Assemble CBX8-BCOR Complex to Repress Bivalent Promoters, Mediate Germinal Center Formation and Lymphomagenesis. Cancer Cell 30, 197–213. - PMC - PubMed
-
- Chen J, Zhang K, Xu Y, Gao Y, Li C, Wang R, and Chen L (2016). The role of microRNA-26a in human cancer progression and clinical application. Tumour Biol 37, 7095–7108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
