

The Selaginella rhizophore has a unique transcriptional identity compared with root and shoot meristems
- PMID: 30614003
- PMCID: PMC6559876
- DOI: 10.1111/nph.15630
The Selaginella rhizophore has a unique transcriptional identity compared with root and shoot meristems
Abstract
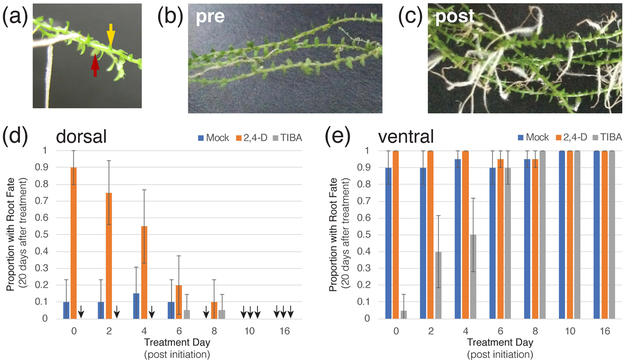
The genus Selaginella resides in an early branch of the land plant lineage that possesses a vasculature and roots. The majority of the Selaginella root system is shoot borne and emerges through a distinctive structure known as the rhizophore, the organ identity of which has been a long-debated question. The rhizophore of Selaginella moellendorffii - a model for the lycophytes - shows plasticity to develop into a root or shoot up until 8 d after angle meristem emergence, after which it is committed to root fate. We subsequently use morphology and plasticity to define the stage of rhizophore identity. Transcriptomic analysis of the rhizophore during its plastic stage reveals that, despite some resemblance to the root meristem, rhizophore gene expression patterns are largely distinct from both shoot and root meristems. Based on this transcriptomic analysis and on historical anatomical work, we conclude that the rhizophore is a distinct organ with unique features.
Keywords: Selaginella moellendorffii; early meristem; lycophyte evolution; lycophyte roots; rhizophore.
© 2018 The Authors. New Phytologist © 2018 New Phytologist Trust.
Figures





References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van Der Straeten D, Peng J, Harberd NP. 2006. Integration of plant responses to environmentally activated phytohormonal signals. Science 311(5757): 91–94. - PubMed
-
- Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, Galinha C, Nussaume L, Noh Y-S, Amasino R, Scheres B. 2004. The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell 119(1): 109–120. - PubMed
-
- Banks JA. 2009. Selaginella and 400 million years of separation. Annu Rev Plant Biol 60: 223–238. - PubMed
-
- Banks JA. 2015. The evolution of the shoot apical meristem from a gene expression perspective. New Phytol 207(3): 486–487. - PubMed
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
