

Circadian rhythm of redox state regulates membrane excitability in hippocampal CA1 neurons
- PMID: 30614107
- PMCID: PMC6609501
- DOI: 10.1111/ejn.14334
Circadian rhythm of redox state regulates membrane excitability in hippocampal CA1 neurons
Abstract
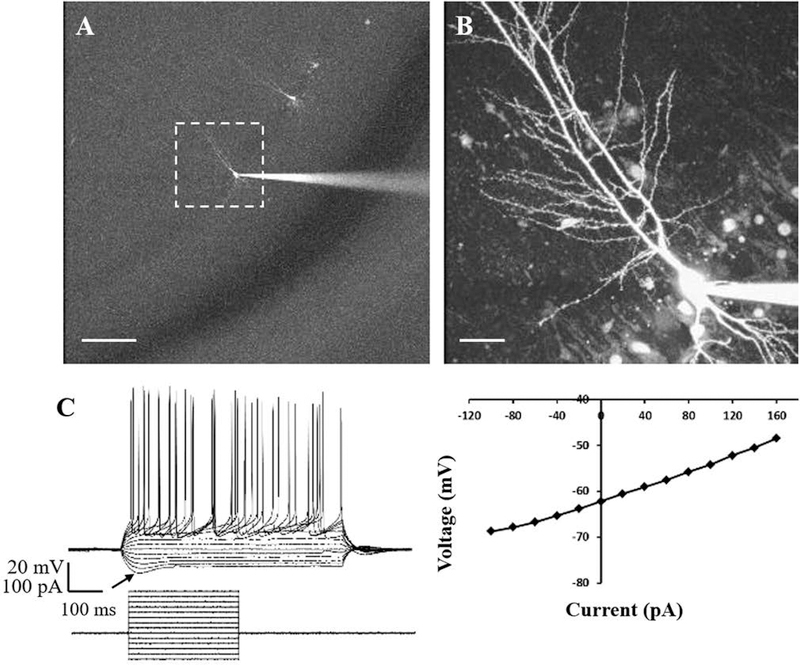
Behaviors, such as sleeping, foraging, and learning, are controlled by different regions of the rat brain, yet they occur rhythmically over the course of day and night. They are aligned adaptively with the day-night cycle by an endogenous circadian clock in the suprachiasmatic nucleus (SCN), but local mechanisms of rhythmic control are not established. The SCN expresses a ~24-hr oscillation in reduction-oxidation that modulates its own neuronal excitability. Could circadian redox oscillations control neuronal excitability elsewhere in the brain? We focused on the CA1 region of the rat hippocampus, which is known for integrating information as memories and where clock gene expression undergoes a circadian oscillation that is in anti-phase to the SCN. Evaluating long-term imaging of endogenous redox couples and biochemical determination of glutathiolation levels, we observed oscillations with a ~24 hr period that is 180° out-of-phase to the SCN. Excitability of CA1 pyramidal neurons, primary hippocampal projection neurons, also exhibits a rhythm in resting membrane potential that is circadian time-dependent and opposite from that of the SCN. The reducing reagent glutathione rapidly and reversibly depolarized the resting membrane potential of CA1 neurons; the magnitude is time-of-day-dependent and, again, opposite from the SCN. These findings extend circadian redox regulation of neuronal excitability from the SCN to the hippocampus. Insights into this system contribute to understanding hippocampal circadian processes, such as learning and memory, seizure susceptibility, and memory loss with aging.
Keywords: CA1 pyramidal neurons; circadian clock; rat hippocampus; reduction-oxidation; suprachiasmatic nucleus.
© 2019 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Conflict of interest statement
COMPETING INTERESTS
The authors have no conflict of interest to declare.
Figures





References
-
- Andersen P, Bliss TV, Lomo T, Olsen LI, Skrede KK (1969) Lamellar organization of hippocampal excitatory pathways. Acta. Physiol. Scand, 76: 4A–5A. - PubMed
Publication types
MeSH terms
Grants and funding
- Beckman Institute Graduate Fellows Program, University of Illinois/International
- U01 MH109062/MH/NIMH NIH HHS/United States
- IGERT CMMB 0965918/National Science Foundation, Division of Graduate Education/International
- R21 MH117377/MH/NIMH NIH HHS/United States
- STC EBICS 0939511/National Science Foundation, Division of Chemical, Bioengineering, Environmental, and Transport Systems/International
LinkOut - more resources
Full Text Sources
Miscellaneous
