

Disruption of the hippocampal and hypothalamic blood-brain barrier in a diet-induced obese model of type II diabetes: prevention and treatment by the mitochondrial carbonic anhydrase inhibitor, topiramate
- PMID: 30616618
- PMCID: PMC6323732
- DOI: 10.1186/s12987-018-0121-6
Disruption of the hippocampal and hypothalamic blood-brain barrier in a diet-induced obese model of type II diabetes: prevention and treatment by the mitochondrial carbonic anhydrase inhibitor, topiramate
Abstract
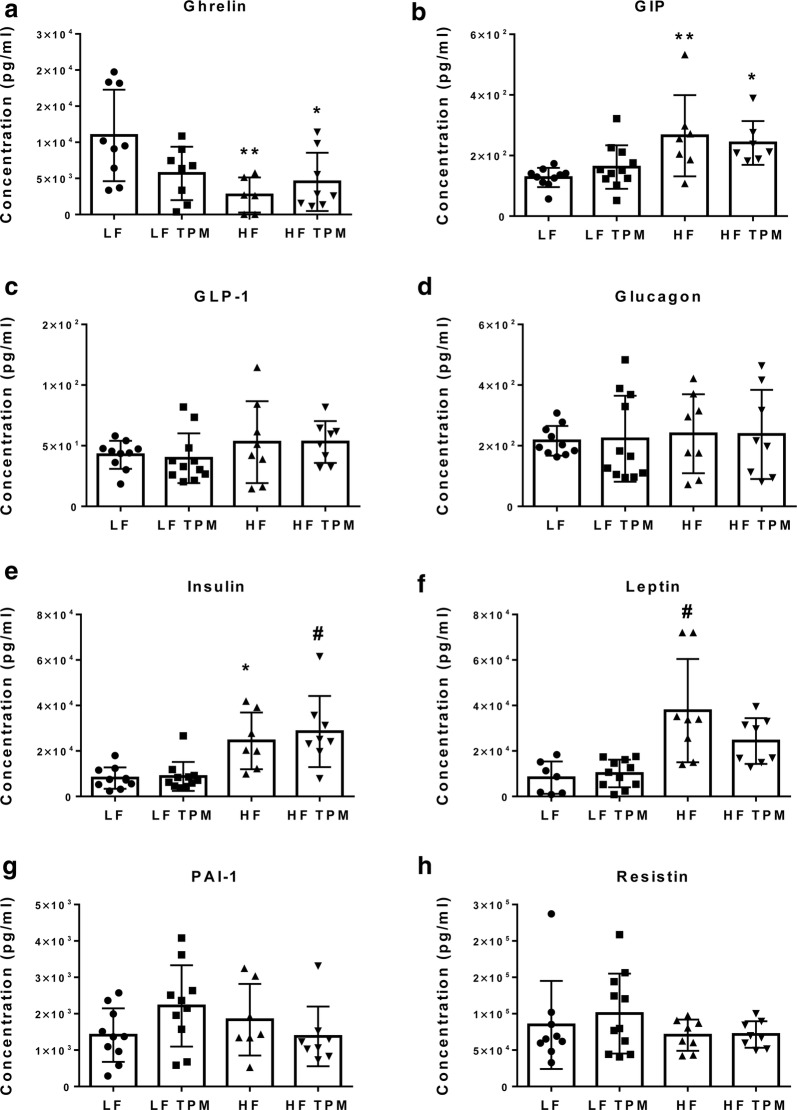
Background: Type II diabetes is a vascular risk factor for cognitive impairment and increased risk of dementia. Disruption of the blood-retinal barrier (BRB) and blood-brain barrier (BBB) are hallmarks of subsequent retinal edema and central nervous system dysfunction. However, the mechanisms by which diet or metabolic syndrome induces dysfunction are not understood. A proposed mechanism is an increase in reactive oxygen species (ROS) and oxidative stress. Inhibition of mitochondrial carbonic anhydrase (mCA) decreases ROS and oxidative stress. In this study, topiramate, a mCA inhibitor, was examined for its ability to protect the BRB and BBB in diet-induced obese type II diabetic mice.
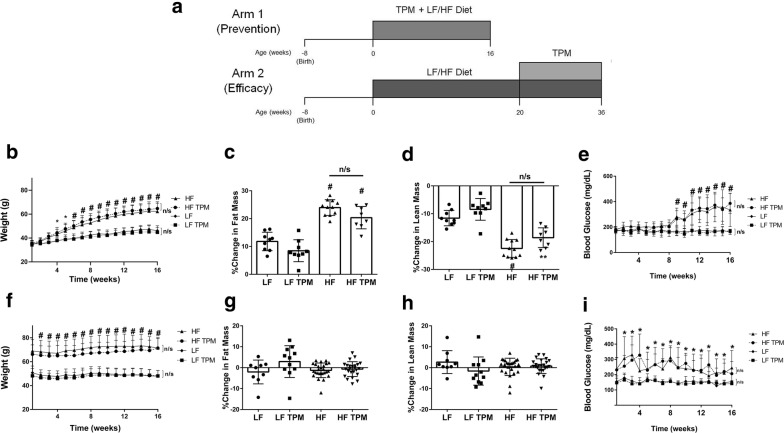
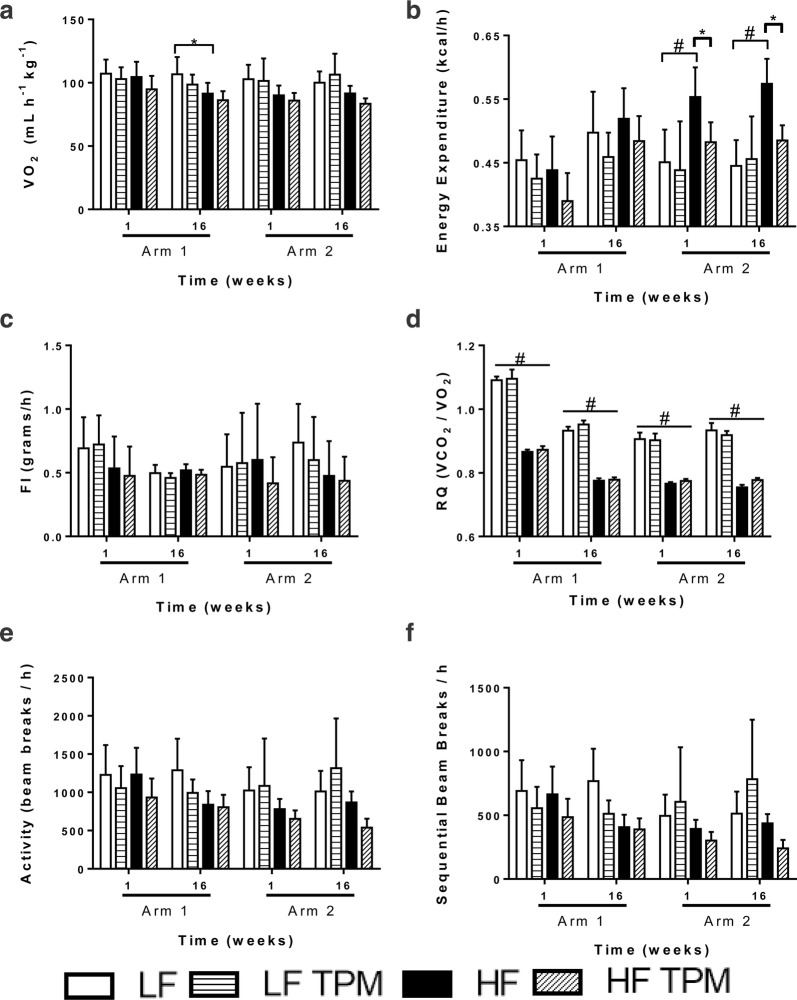
Methods: BBB and BRB permeability were assessed using 14C-sucrose and 99mTc-albumin in CD-1 mice fed a low-fat (control) or a high-fat diet. Topiramate administration was compared to saline controls in both preventative and efficacy arms examining BRB and BBB disruption. Body weight and blood glucose were measured weekly and body composition was assessed using EchoMRI. Metabolic activity was measured using a comprehensive laboratory animal monitoring system. Brain tissues collected from the mice were assessed for changes in oxidative stress and tight junction proteins.
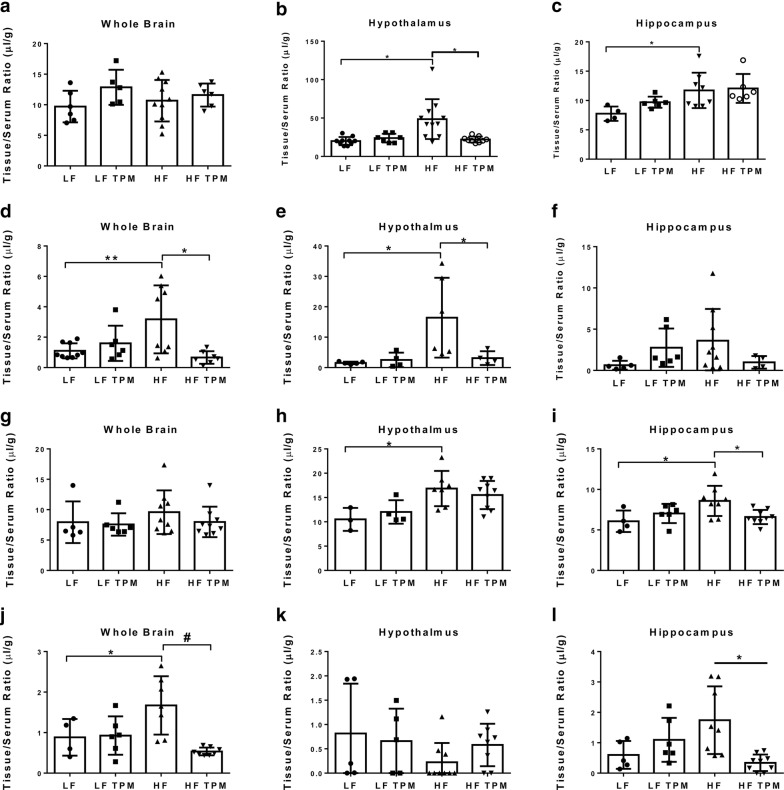
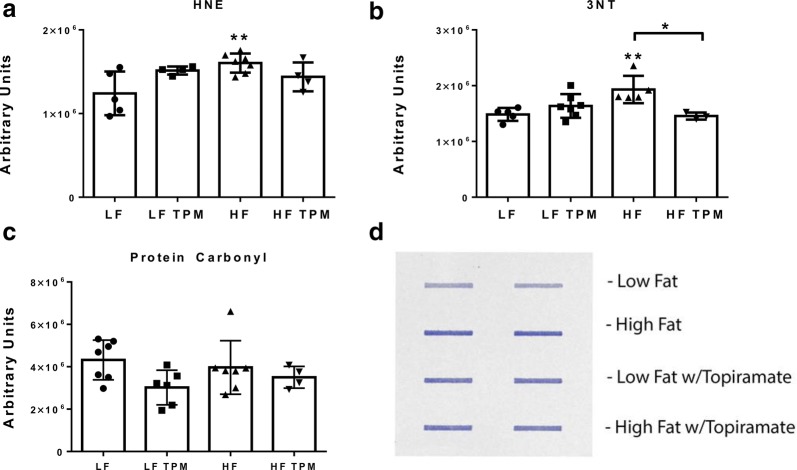
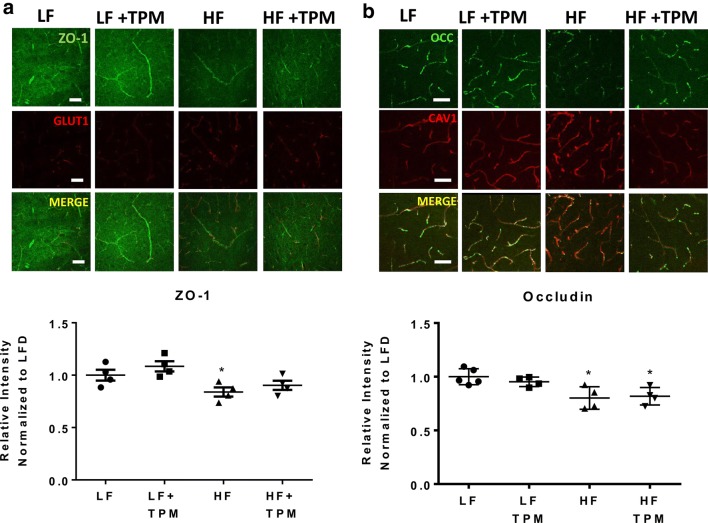
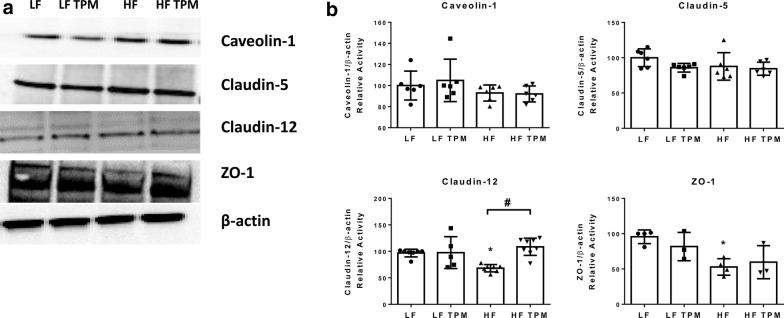
Results: High-fat feeding caused increased entry of 14C-sucrose and 99mTc-albumin into the brains of diet-induced obese type II diabetic mice. Increased permeability to 14C-sucrose was observed in the hypothalamus and hippocampus, and attenuated by topiramate treatment, while increased permeability to 99mTc-albumin occurred in the whole brain and was also attenuated by topiramate. Treatment with topiramate decreased measures of oxidative stress and increased expression of the tight junction proteins ZO-1 and claudin-12. In the retina, we observed increased entry of 99mTc-albumin simultaneously with increased entry into the whole brain during the preventative arm. This occurred prior to increased entry to the retina for 14C-sucrose which occurred during the efficacy arm. Treatment with topiramate had no effect on the retina.
Conclusions: Blood-brain barrier and blood-retinal barrier dysfunction were examined in a mouse model of diet-induced obese type II diabetes. These studies demonstrate that there are spatial and temporal differences in 14C-sucrose and 99mTc-albumin permeability in the brain and retina of diet-induced obese type II diabetic mice. Topiramate, a mitochondrial carbonic anhydrase inhibitor, is efficacious at both preventing and treating BBB disruption in this diet-induced obese type II diabetic mouse model.
Keywords: Blood–brain barrier; Blood–retinal barrier; Hippocampus; Hypothalamus; Topiramate; Type II diabetes.
Figures
References
-
- Ozkan E, Gocmen R, Topcuoglu MA, Arsava EM. Blood–retina-barrier disruption accompanying blood–brain-barrier dysfunction in posterior reversible encephalopathy syndrome. J Neurol Sci. 2014;346(1):315–317. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
