

Understanding phosphoinositides: rare, dynamic, and essential membrane phospholipids
- PMID: 30617162
- PMCID: PMC6342281
- DOI: 10.1042/BCJ20180022
Understanding phosphoinositides: rare, dynamic, and essential membrane phospholipids
Abstract
Polyphosphoinositides (PPIs) are essential phospholipids located in the cytoplasmic leaflet of eukaryotic cell membranes. Despite contributing only a small fraction to the bulk of cellular phospholipids, they make remarkable contributions to practically all aspects of a cell's life and death. They do so by recruiting cytoplasmic proteins/effectors or by interacting with cytoplasmic domains of membrane proteins at the membrane-cytoplasm interface to organize and mold organelle identity. The present study summarizes aspects of our current understanding concerning the metabolism, manipulation, measurement, and intimate roles these lipids play in regulating membrane homeostasis and vital cell signaling reactions in health and disease.
Keywords: PTEN; lipid transfer; phosphatidylinositol; phosphoinositide 3-kinase; transmembrane proteins.
© 2019 The Author(s). Published by Portland Press Limited on behalf of the Biochemical Society.
Conflict of interest statement
Competing Interests
The Authors declare that there are no competing interests associated with the manuscript.
Figures
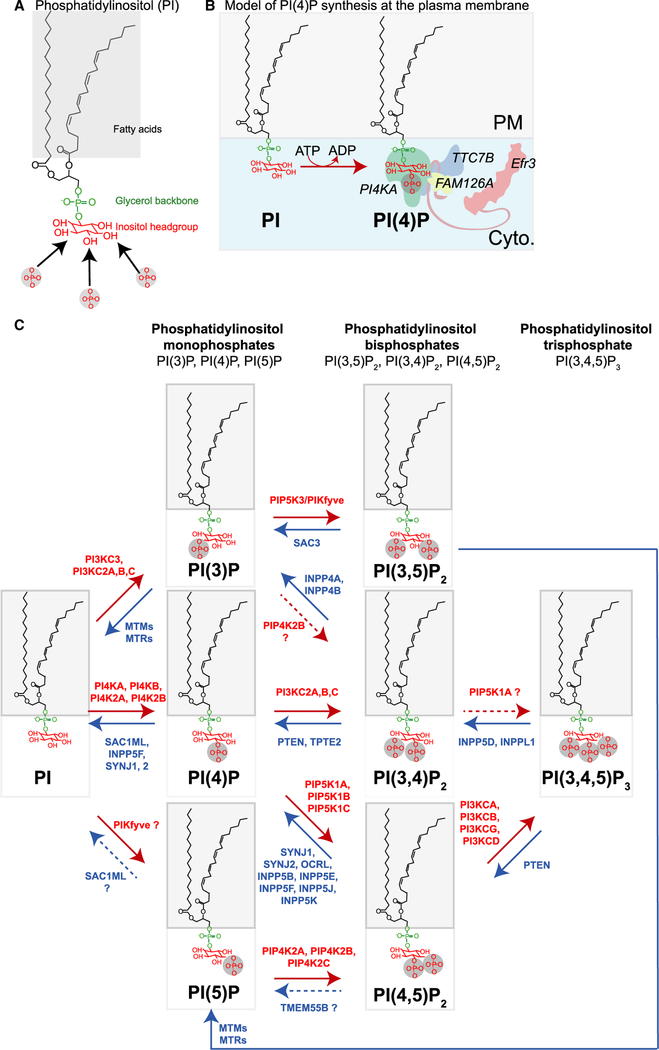
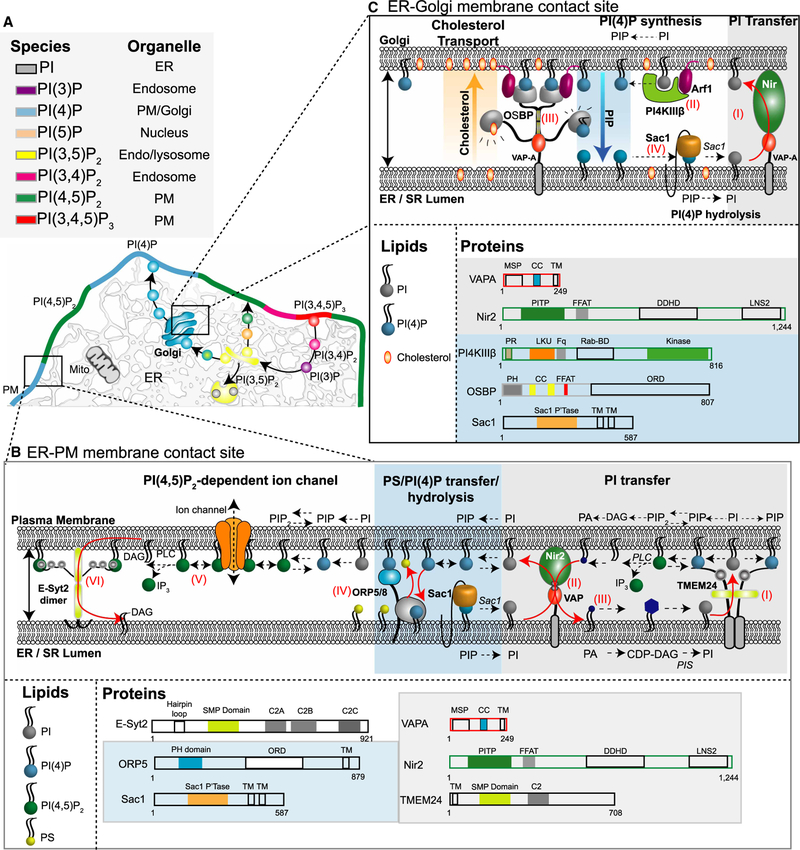
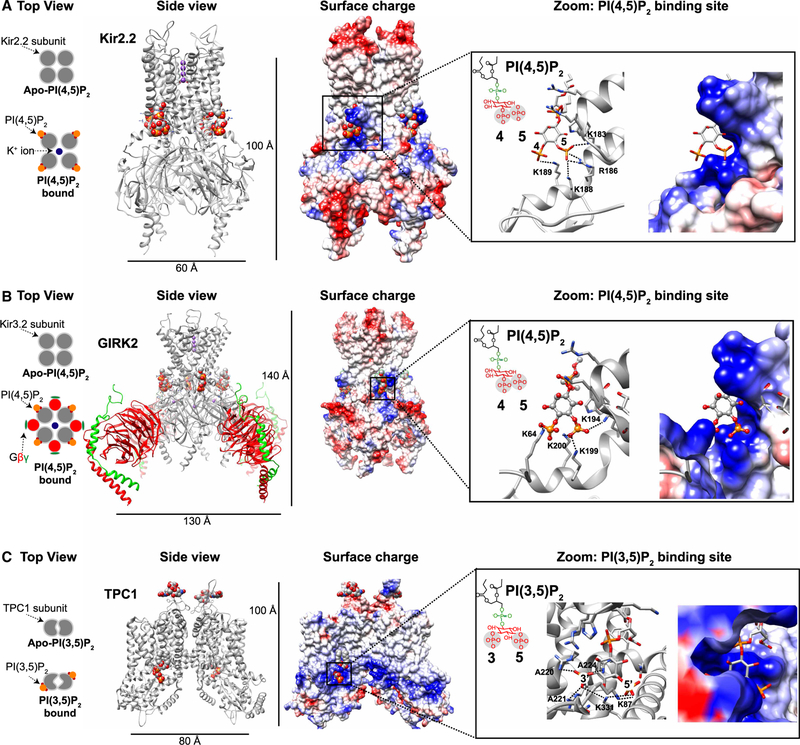
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials