

A Hippocampal Model for Behavioral Time Acquisition and Fast Bidirectional Replay of Spatio-Temporal Memory Sequences
- PMID: 30618583
- PMCID: PMC6306028
- DOI: 10.3389/fnins.2018.00961
A Hippocampal Model for Behavioral Time Acquisition and Fast Bidirectional Replay of Spatio-Temporal Memory Sequences
Abstract
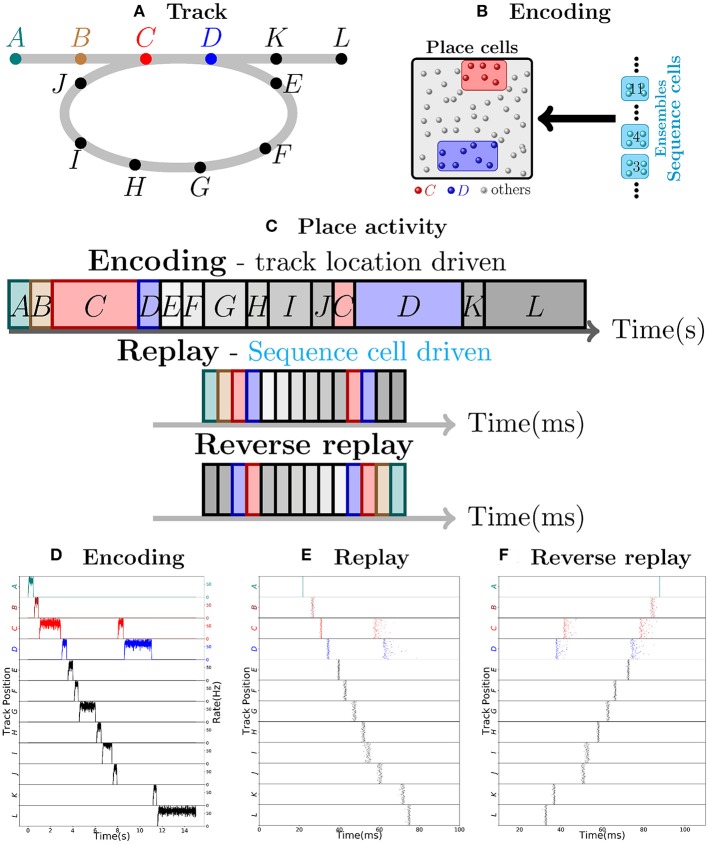
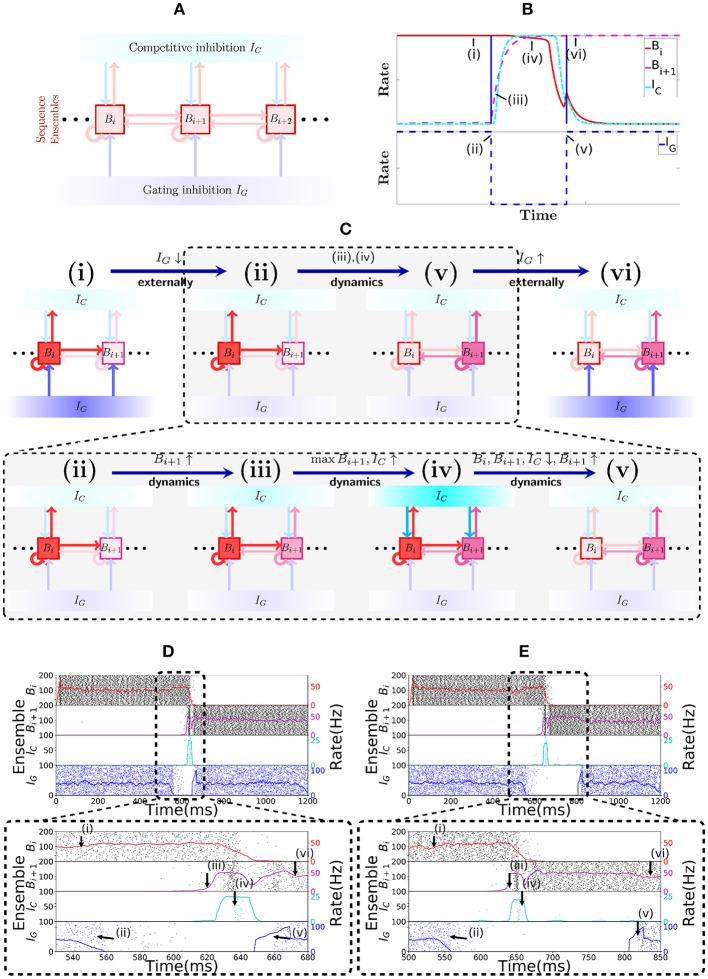
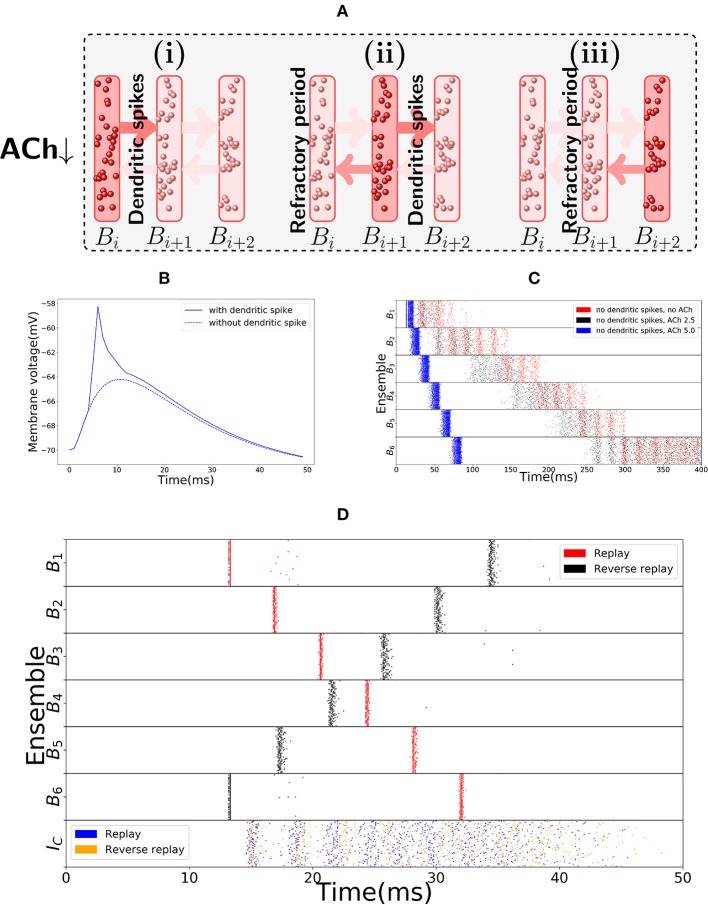
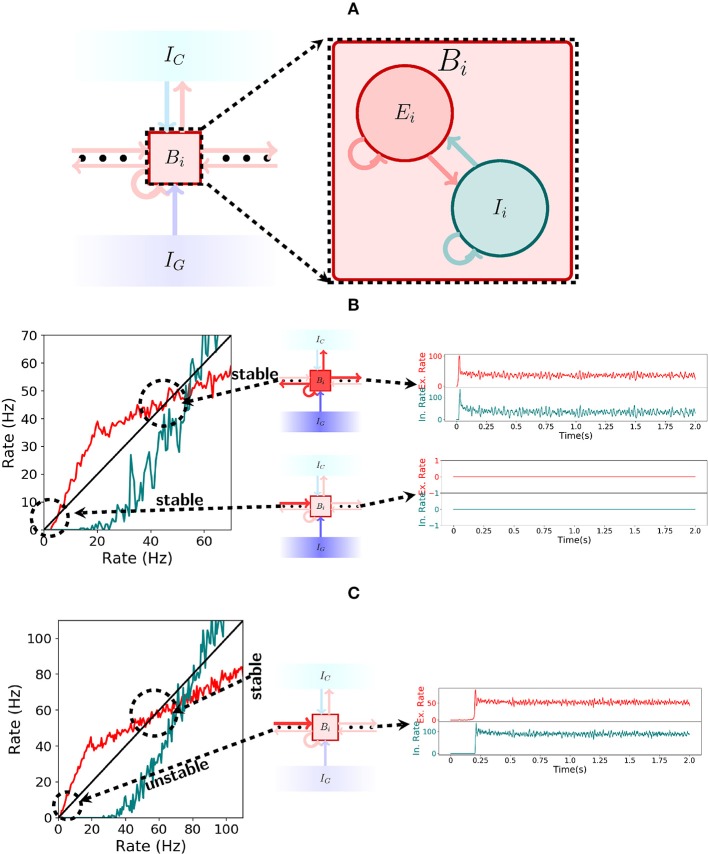
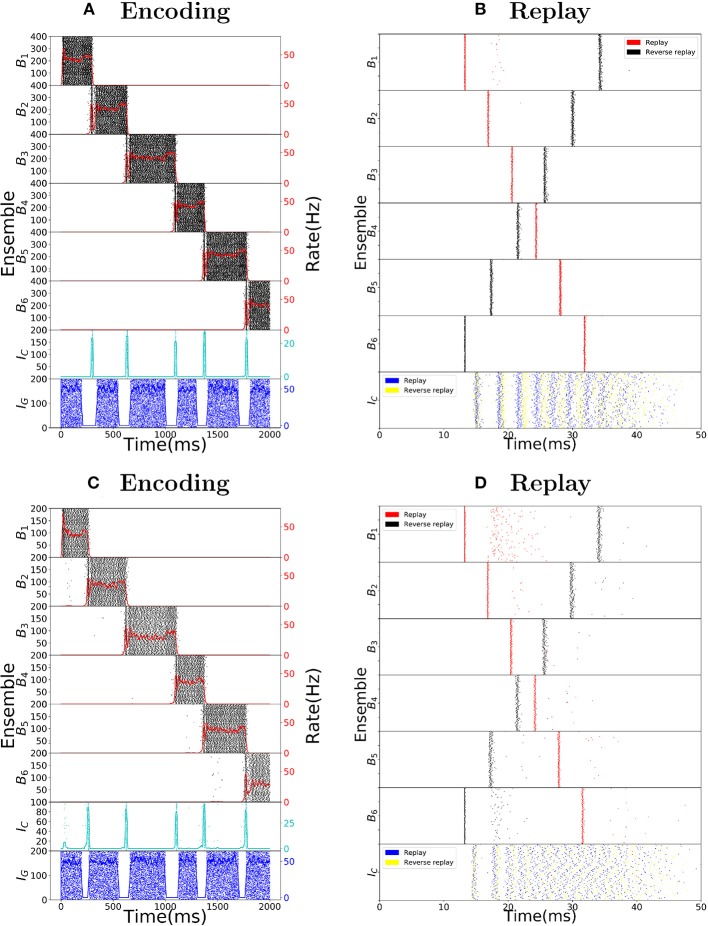
The hippocampus is known to play a crucial role in the formation of long-term memory. For this, fast replays of previously experienced activities during sleep or after reward experiences are believed to be crucial. But how such replays are generated is still completely unclear. In this paper we propose a possible mechanism for this: we present a model that can store experienced trajectories on a behavioral timescale after a single run, and can subsequently bidirectionally replay such trajectories, thereby omitting any specifics of the previous behavior like speed, etc, but allowing repetitions of events, even with different subsequent events. Our solution builds on well-known concepts, one-shot learning and synfire chains, enhancing them by additional mechanisms using global inhibition and disinhibition. For replays our approach relies on dendritic spikes and cholinergic modulation, as supported by experimental data. We also hypothesize a functional role of disinhibition as a pacemaker during behavioral time.
Keywords: cholinergic modulation; dendritic spikes; disinhibition; hippocampal replays; one-shot learning; place cells; reverse replays; synfire chains.
Figures
References
-
- Abeles M. (1982). Local Cortical Circuits. Berlin: Springer.
-
- Amit D. J., Fusi S. (1994). Learning in neural networks with material synapses. Neural Comput. 6, 957–982. 10.1162/neco.1994.6.5.957 - DOI
LinkOut - more resources
Full Text Sources