

The Complex Conformational Dynamics of Neuronal Calcium Sensor-1: A Single Molecule Perspective
- PMID: 30618617
- PMCID: PMC6304440
- DOI: 10.3389/fnmol.2018.00468
The Complex Conformational Dynamics of Neuronal Calcium Sensor-1: A Single Molecule Perspective
Abstract
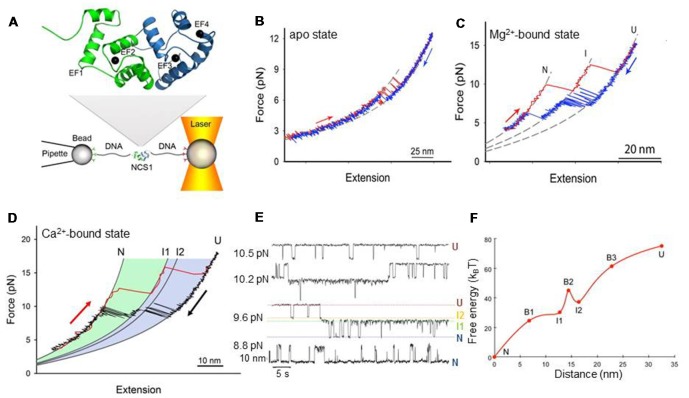
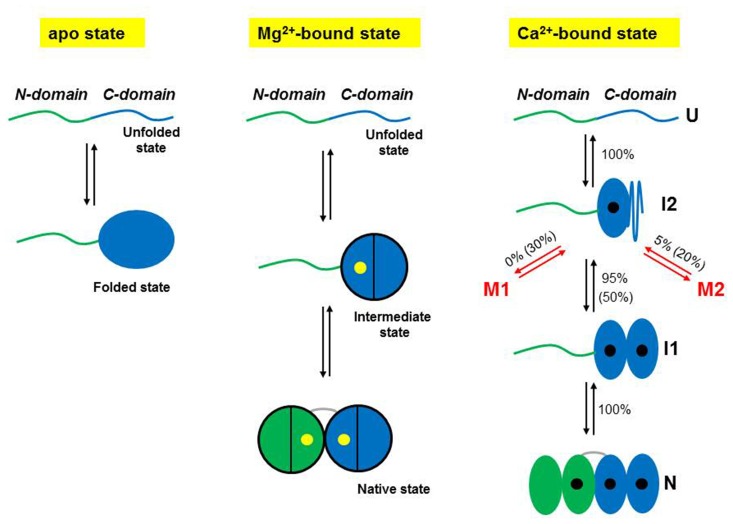
The human neuronal calcium sensor-1 (NCS-1) is a multispecific two-domain EF-hand protein expressed predominantly in neurons and is a member of the NCS protein family. Structure-function relationships of NCS-1 have been extensively studied showing that conformational dynamics linked to diverse ion-binding is important to its function. NCS-1 transduces Ca2+ changes in neurons and is linked to a wide range of neuronal functions such as regulation of neurotransmitter release, voltage-gated Ca2+ channels and neuronal outgrowth. Defective NCS-1 can be deleterious to cells and has been linked to serious neuronal disorders like autism. Here, we review recent studies describing at the single molecule level the structural and mechanistic details of the folding and misfolding processes of the non-myristoylated NCS-1. By manipulating one molecule at a time with optical tweezers, the conformational equilibria of the Ca2+-bound, Mg2+-bound and apo states of NCS-1 were investigated revealing a complex folding mechanism underlain by a rugged and multidimensional energy landscape. The molecular rearrangements that NCS-1 undergoes to transit from one conformation to another and the energetics of these reactions are tightly regulated by the binding of divalent ions (Ca2+ and Mg2+) to its EF-hands. At pathologically high Ca2+ concentrations the protein sometimes follows non-productive misfolding pathways leading to kinetically trapped and potentially harmful misfolded conformations. We discuss the significance of these misfolding events as well as the role of inter-domain interactions in shaping the energy landscape and ultimately the biological function of NCS-1. The conformational equilibria of NCS-1 are also compared to those of calmodulin (CaM) and differences and similarities in the behavior of these proteins are rationalized in terms of structural properties.
Keywords: NCS-1; calcium binding; optical tweezers; protein folding and misfolding; single molecule studies.
Figures
References
-
- Aravind P., Chandra K., Reddy P. P., Jeromin A., Chary K., Sharma Y. (2008). Regulatory and structural EF-hand motifs of neuronal calcium sensor-1: Mg2+ modulates Ca2+ binding, Ca2+-induced conformational changes, and equilibrium unfolding transitions. J. Mol. Biol. 376, 1100–1115. 10.1016/j.jmb.2007.12.033 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous