

Relevance of Bacteriophage 933W in the Development of Hemolytic Uremic Syndrome (HUS)
- PMID: 30619183
- PMCID: PMC6300567
- DOI: 10.3389/fmicb.2018.03104
Relevance of Bacteriophage 933W in the Development of Hemolytic Uremic Syndrome (HUS)
Abstract
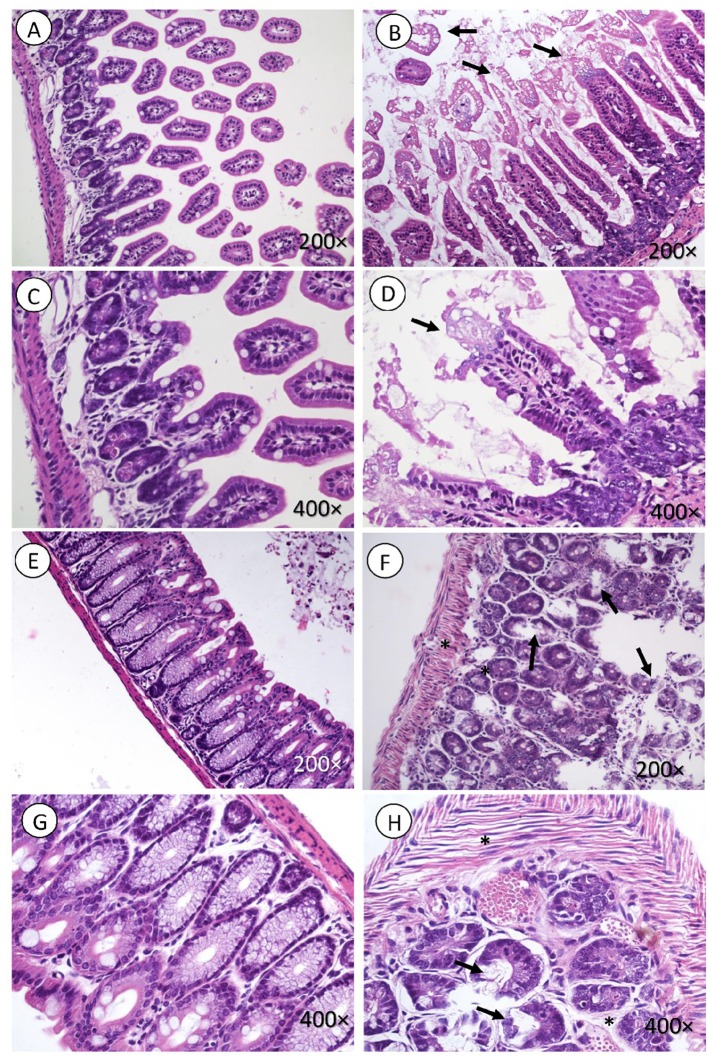
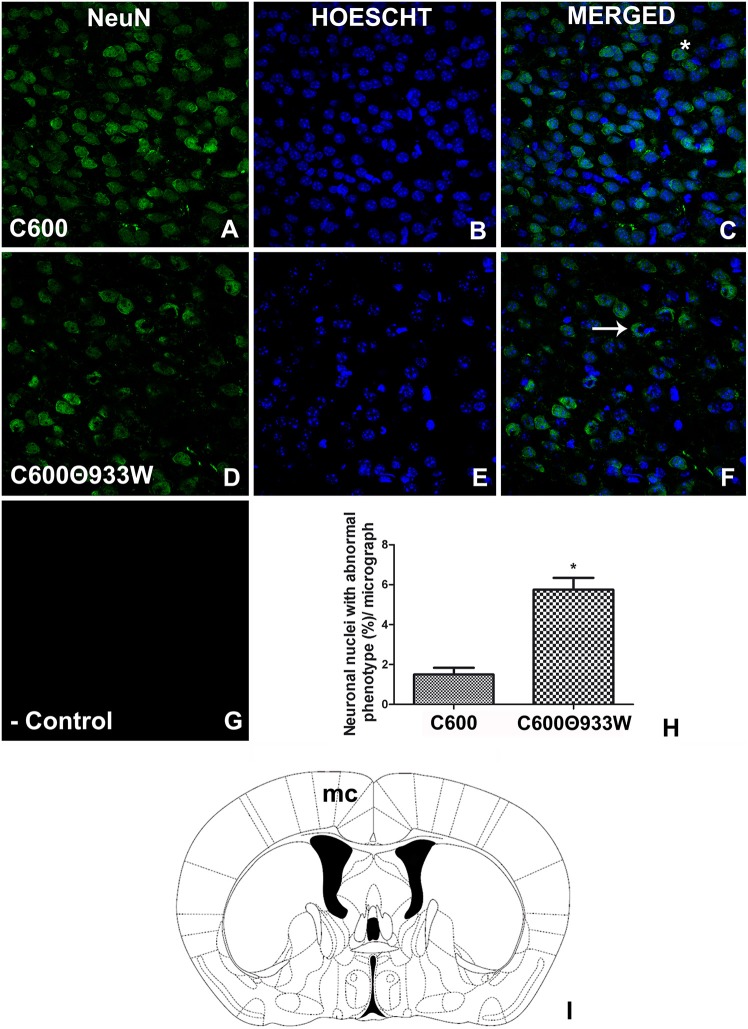
Hemolytic uremic syndrome (HUS), principally caused by shiga toxins (Stxs), is associated with Shiga toxin-producing Escherichia coli (STEC) infections. We previously reported Stx2 expression by host cells in vitro and in vivo. As the genes encoding the two Stx subunits are located in bacteriophage genomes, the aim of the current study was to evaluate the role of bacteriophage induction in HUS development in absence of an E. coli O157:H7 genomic background. Mice were inoculated with a non-pathogenic E. coli strain carrying the lysogenic bacteriophage 933W (C600Φ933W), and bacteriophage excision was induced by an antibiotic. The mice died 72 h after inoculation, having developed pathogenic damage typical of STEC infection. As well as renal and intestinal damage, markers of central nervous system (CNS) injury were observed, including aberrant immunolocalization of neuronal nuclei (NeuN) and increased expression of glial fibrillary acidic protein (GFAP). These results show that bacteriophage 933W without an E. coli O157:H7 background is capable of inducing the pathogenic damage associated with STEC infection. In addition, a novel mouse model was developed to evaluate therapeutic approaches focused on the bacteriophage as a new target.
Keywords: Shiga toxigenic E. coli (STEC); Shiga toxin (Stx); animal model; bacterio(phages); hemolytic uremic syndrome.
Figures





References
-
- Fernandez-Brando R. J., Cabrera G., Baschkíer A., Mejías M. P., Panek C. A., Miliwebsky E., et al. . (2014). Oral administration of Shiga toxin-producing Escherichia coli induces intestinal and systemic specific immune response in mice. Med. Microbiol. Immunol. 203, 145–54. 10.1007/s00430-013-0325-y - DOI - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous