

Iron Regulation in Clostridioides difficile
- PMID: 30619231
- PMCID: PMC6311696
- DOI: 10.3389/fmicb.2018.03183
Iron Regulation in Clostridioides difficile
Abstract
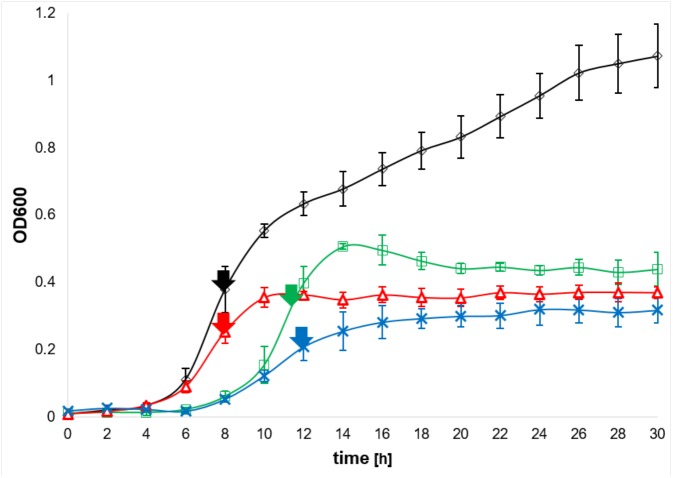
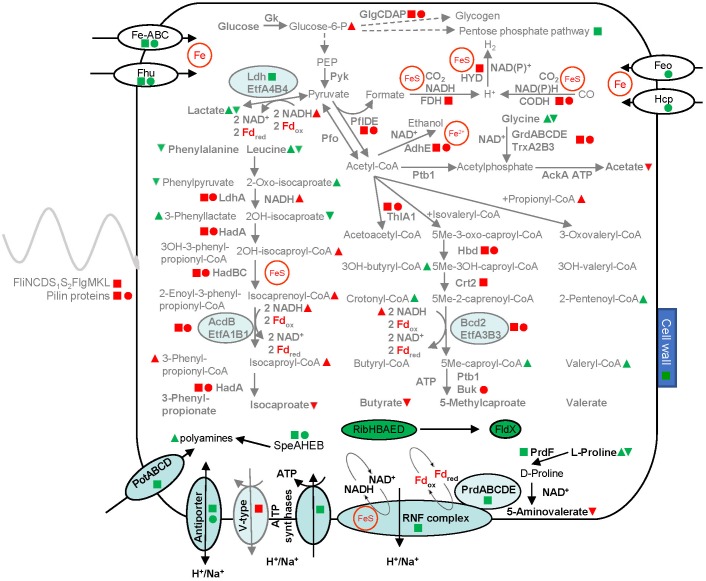
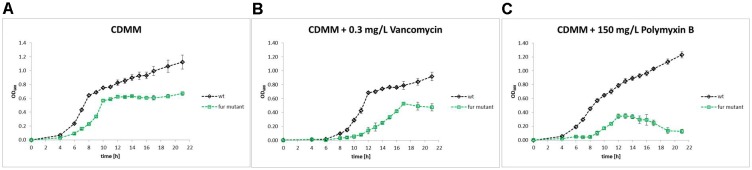
The response to iron limitation of several bacteria is regulated by the ferric uptake regulator (Fur). The Fur-regulated transcriptional, translational and metabolic networks of the Gram-positive, pathogen Clostridioides difficile were investigated by a combined RNA sequencing, proteomic, metabolomic and electron microscopy approach. At high iron conditions (15 μM) the C. difficile fur mutant displayed a growth deficiency compared to wild type C. difficile cells. Several iron and siderophore transporter genes were induced by Fur during low iron (0.2 μM) conditions. The major adaptation to low iron conditions was observed for the central energy metabolism. Most ferredoxin-dependent amino acid fermentations were significantly down regulated (had, etf, acd, grd, trx, bdc, hbd). The substrates of these pathways phenylalanine, leucine, glycine and some intermediates (phenylpyruvate, 2-oxo-isocaproate, 3-hydroxy-butyryl-CoA, crotonyl-CoA) accumulated, while end products like isocaproate and butyrate were found reduced. Flavodoxin (fldX) formation and riboflavin biosynthesis (rib) were enhanced, most likely to replace the missing ferredoxins. Proline reductase (prd), the corresponding ion pumping RNF complex (rnf) and the reaction product 5-aminovalerate were significantly enhanced. An ATP forming ATPase (atpCDGAHFEB) of the F0F1-type was induced while the formation of a ATP-consuming, proton-pumping V-type ATPase (atpDBAFCEKI) was decreased. The [Fe-S] enzyme-dependent pyruvate formate lyase (pfl), formate dehydrogenase (fdh) and hydrogenase (hyd) branch of glucose utilization and glycogen biosynthesis (glg) were significantly reduced, leading to an accumulation of glucose and pyruvate. The formation of [Fe-S] enzyme carbon monoxide dehydrogenase (coo) was inhibited. The fur mutant showed an increased sensitivity to vancomycin and polymyxin B. An intensive remodeling of the cell wall was observed, Polyamine biosynthesis (spe) was induced leading to an accumulation of spermine, spermidine, and putrescine. The fur mutant lost most of its flagella and motility. Finally, the CRISPR/Cas and a prophage encoding operon were downregulated. Fur binding sites were found upstream of around 20 of the regulated genes. Overall, adaptation to low iron conditions in C. difficile focused on an increase of iron import, a significant replacement of iron requiring metabolic pathways and the restructuring of the cell surface for protection during the complex adaptation phase and was only partly directly regulated by Fur.
Keywords: Fur; cell wall; iron regulation; iron transport; metabolism; polyamine.
Figures


References
-
- Aboulnaga E.-H., Pinkenburg O., Schiffels J., El-Refai A., Buckel W., Selmer T. (2013). Effect of an oxygen-tolerant bifurcating butyryl coenzyme A dehydrogenase/electron-transferring flavoprotein complex from Clostridium difficile on butyrate production in Escherichia coli. J. Bacteriol. 195 3704–3713. 10.1128/JB.00321-13 - DOI - PMC - PubMed
-
- Argiris Symeonidis and Markos Marangos (2012). “Iron and microbial growth,” in Insight and Control of Infectious Disease in Global Scenario, ed. Roy P. K. (Rijeka: In Tech; ), 289–332.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous