

Parasite Recognition and Signaling Mechanisms in Innate Immune Responses to Malaria
- PMID: 30619355
- PMCID: PMC6305727
- DOI: 10.3389/fimmu.2018.03006
Parasite Recognition and Signaling Mechanisms in Innate Immune Responses to Malaria
Abstract
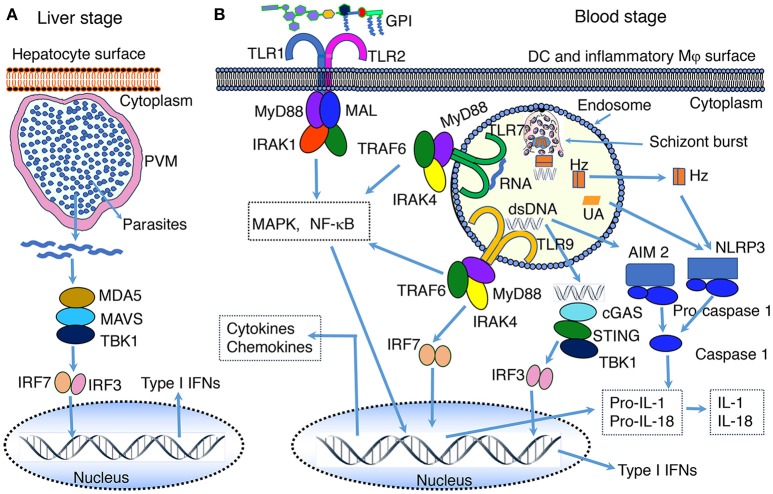
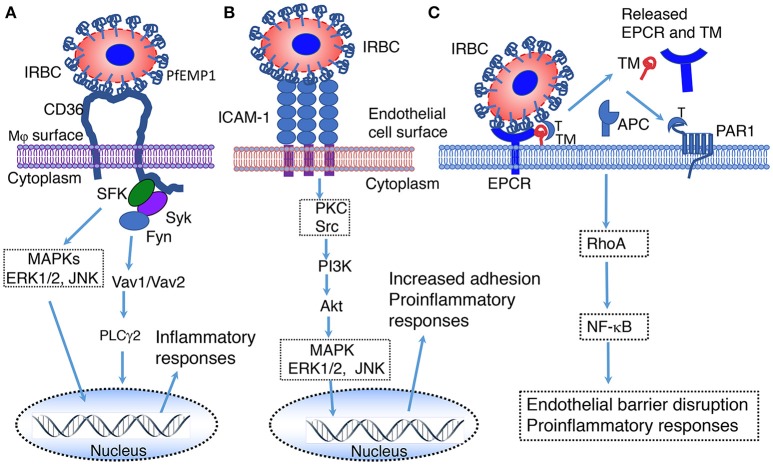
Malaria caused by the Plasmodium family of parasites, especially P.falciparum and P. vivax, is a major health problem in many countries in the tropical and subtropical regions of the world. The disease presents a wide array of systemic clinical conditions and several life-threatening organ pathologies, including the dreaded cerebral malaria. Like many other infectious diseases, malaria is an inflammatory response-driven disease, and positive outcomes to infection depend on finely tuned regulation of immune responses that efficiently clear parasites and allow protective immunity to develop. Immune responses initiated by the innate immune system in response to parasites play key roles both in protective immunity development and pathogenesis. Initial pro-inflammatory responses are essential for clearing infection by promoting appropriate cell-mediated and humoral immunity. However, elevated and prolonged pro-inflammatory responses owing to inappropriate cellular programming contribute to disease conditions. A comprehensive knowledge of the molecular and cellular mechanisms that initiate immune responses and how these responses contribute to protective immunity development or pathogenesis is important for developing effective therapeutics and/or a vaccine. Historically, in efforts to develop a vaccine, immunity to malaria was extensively studied in the context of identifying protective humoral responses, targeting proteins involved in parasite invasion or clearance. The innate immune response was thought to be non-specific. However, during the past two decades, there has been a significant progress in understanding the molecular and cellular mechanisms of host-parasite interactions and the associated signaling in immune responses to malaria. Malaria infection occurs at two stages, initially in the liver through the bite of a mosquito, carrying sporozoites, and subsequently, in the blood through the invasion of red blood cells by merozoites released from the infected hepatocytes. Soon after infection, both the liver and blood stage parasites are sensed by various receptors of the host innate immune system resulting in the activation of signaling pathways and production of cytokines and chemokines. These immune responses play crucial roles in clearing parasites and regulating adaptive immunity. Here, we summarize the knowledge on molecular mechanisms that underlie the innate immune responses to malaria infection.
Keywords: host receptors; immunostimulatory factors; innate immune responses; malaria; pathogenesis; protective immunity; signaling mechanisms.
Figures
References
-
- World Health Organization. World Malaria Report Geneva. (2017). Available online at: http://www.who.int/malaria/publications/world-malaria-report-2017/en/
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous