

A New Role For Green Leaf Volatile Esters in Tomato Stomatal Defense Against Pseudomonas syringe pv. tomato
- PMID: 30619420
- PMCID: PMC6305539
- DOI: 10.3389/fpls.2018.01855
A New Role For Green Leaf Volatile Esters in Tomato Stomatal Defense Against Pseudomonas syringe pv. tomato
Abstract
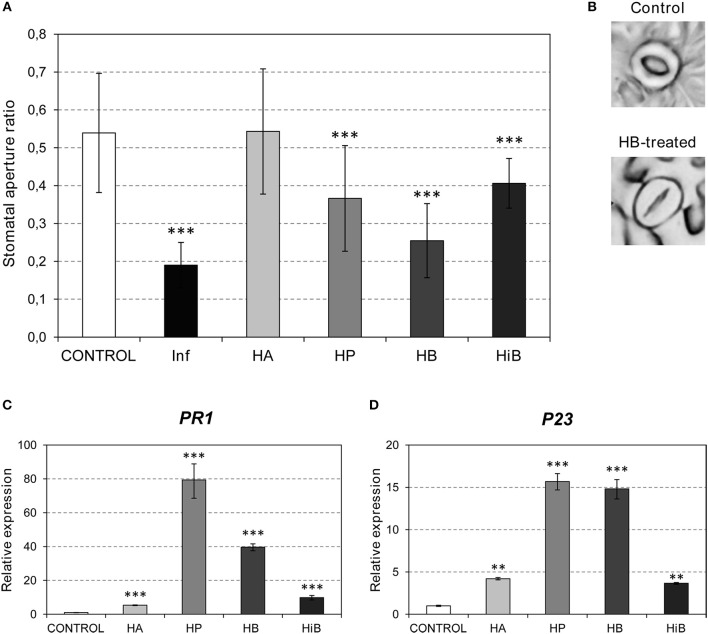
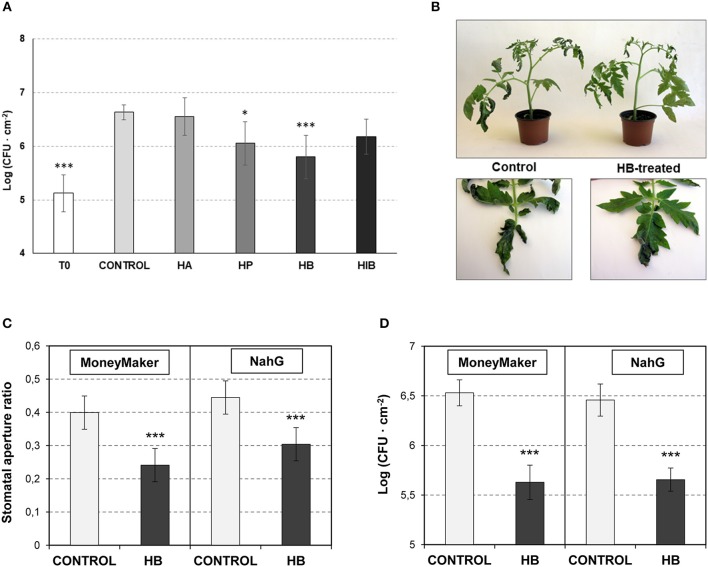
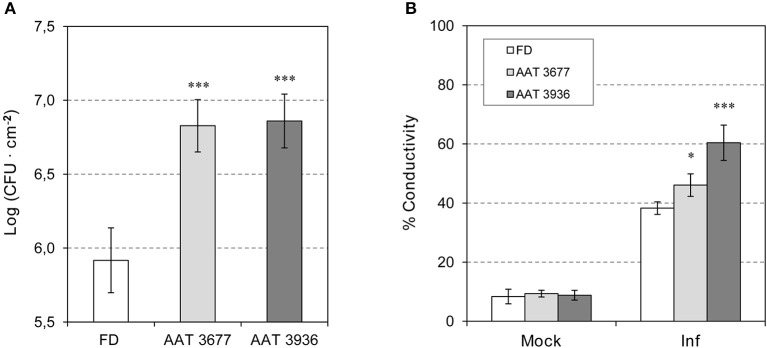
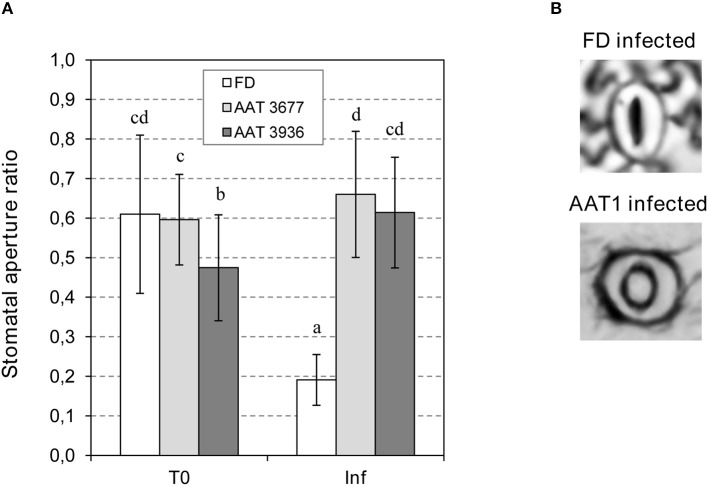
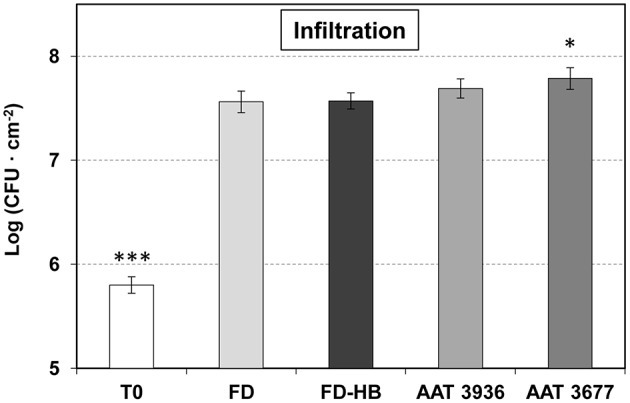
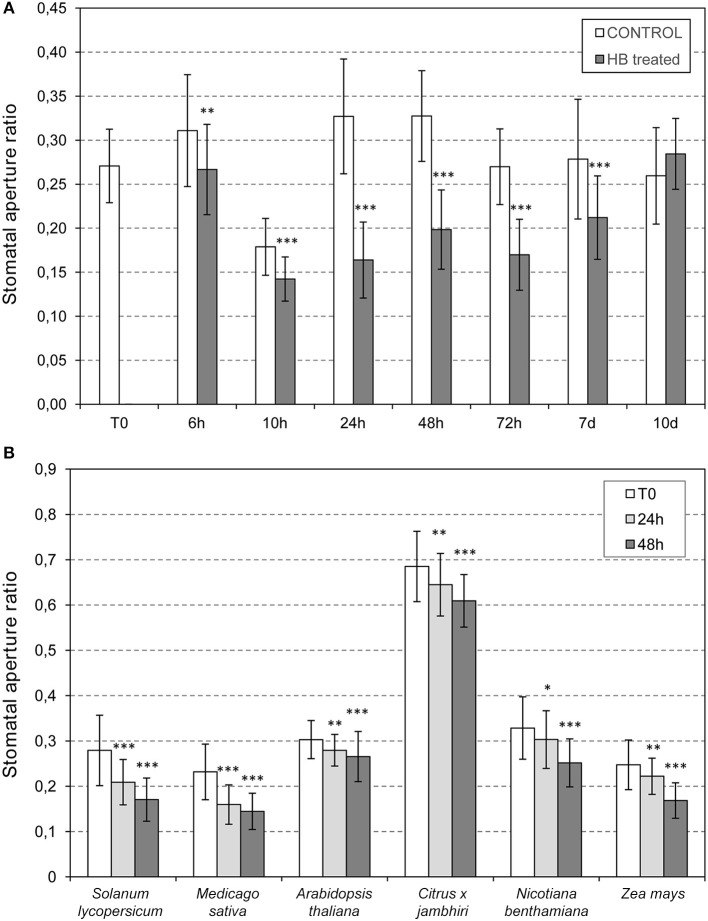
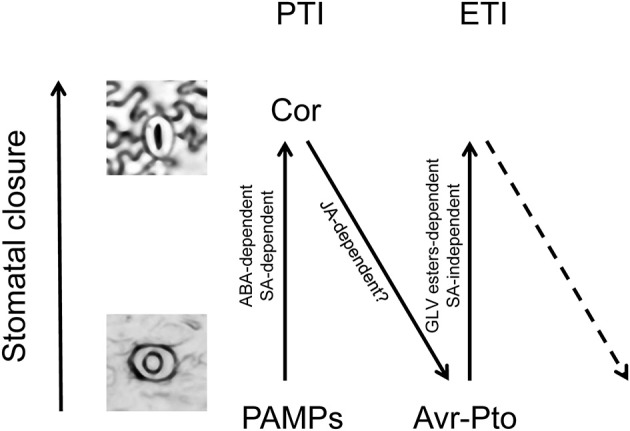
The volatile esters of (Z)-3-hexenol with acetic, propionic, isobutyric, or butyric acids are synthesized by alcohol acyltransferases (AAT) in plants. These compounds are differentially emitted when tomato plants are efficiently resisting an infection with Pseudomonas syringae pv. tomato. We have studied the defensive role of these green leaf volatile (GLV) esters in the tomato response to bacterial infection, by analyzing the induction of resistance mediated by these GLVs and the phenotype upon bacterial infection of tomato plants impaired in their biosynthesis. We observed that treatments of plants with (Z)-3-hexenyl propionate (HP) and, to a greater extent with (Z)-3-hexenyl butyrate (HB), resulted in stomatal closure, PR gene induction and enhanced resistance to the bacteria. HB-mediated stomatal closure was also effective in several plant species belonging to Nicotiana, Arabidopsis, Medicago, Zea and Citrus genus, and both stomatal closure and resistance were induced in HB-treated NahG tomato plants, which are deficient in salicylic acid (SA) accumulation. Transgenic antisense AAT1 tomato plants, which displayed a reduction of ester emissions upon bacterial infection in leaves, exhibited a lower ratio of stomatal closure and were hyper-susceptible to bacterial infection. Our results confirm the role of GLV esters in plant immunity, uncovering a SA-independent effect of HB in stomatal defense. Moreover, we identified HB as a natural stomatal closure compound with potential agricultural applications.
Keywords: AAT1; GLV esters; bacteria; defense; stomata; tomato.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous