

Inward- and outward-facing X-ray crystal structures of homodimeric P-glycoprotein CmABCB1
- PMID: 30622258
- PMCID: PMC6325147
- DOI: 10.1038/s41467-018-08007-x
Inward- and outward-facing X-ray crystal structures of homodimeric P-glycoprotein CmABCB1
Abstract
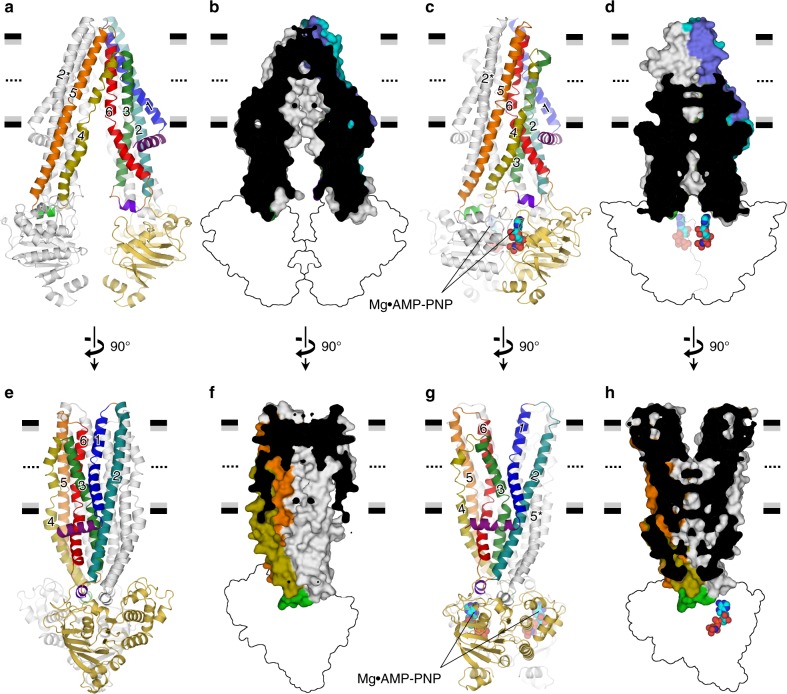
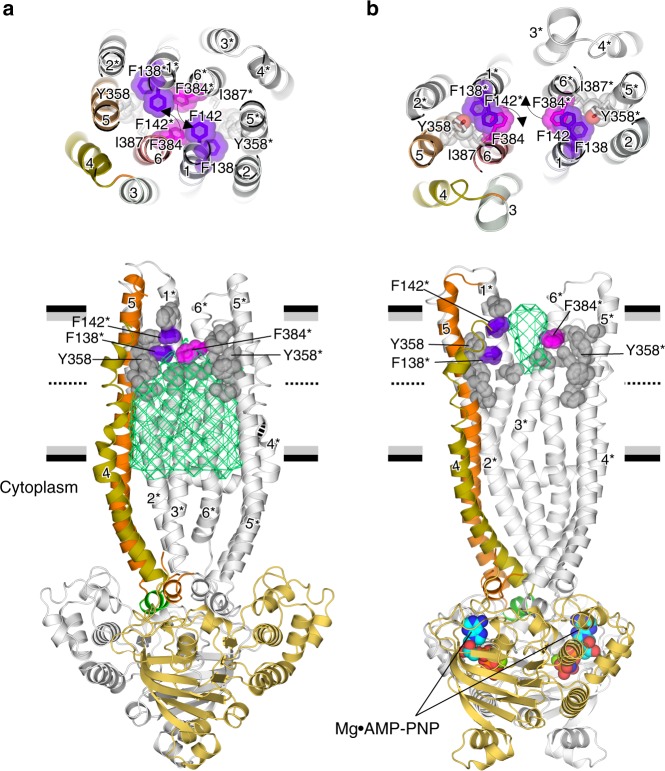
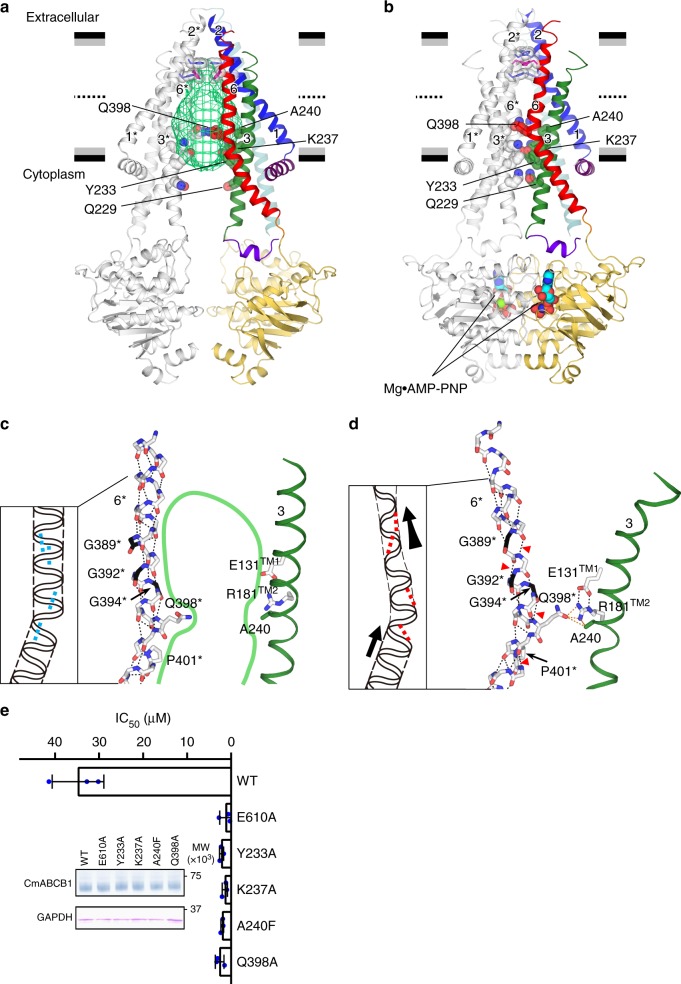
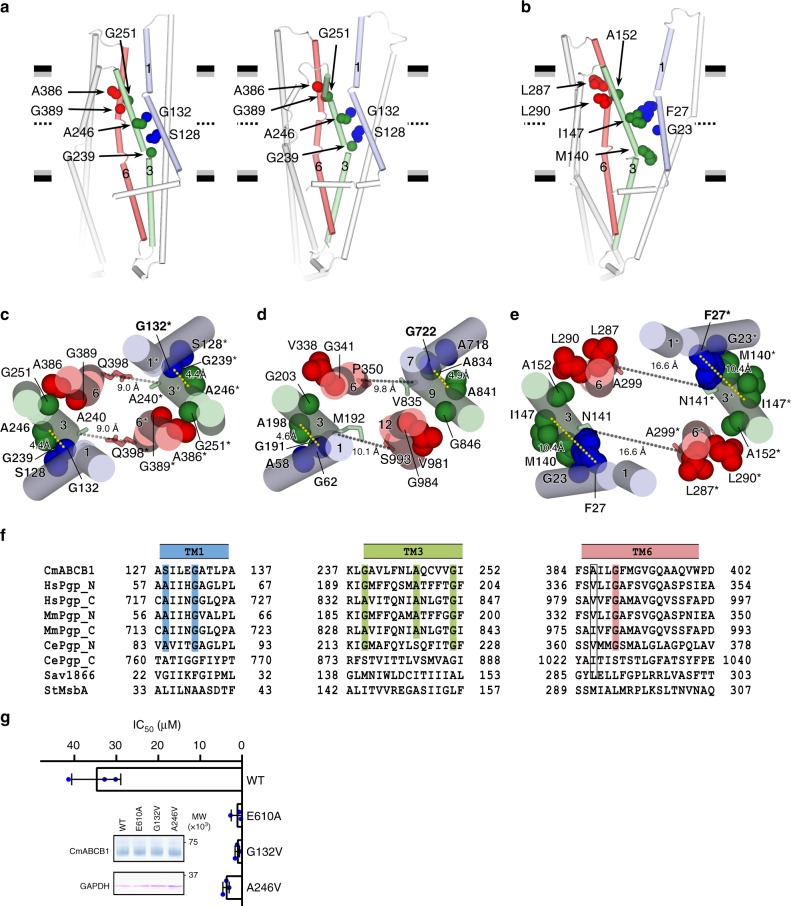
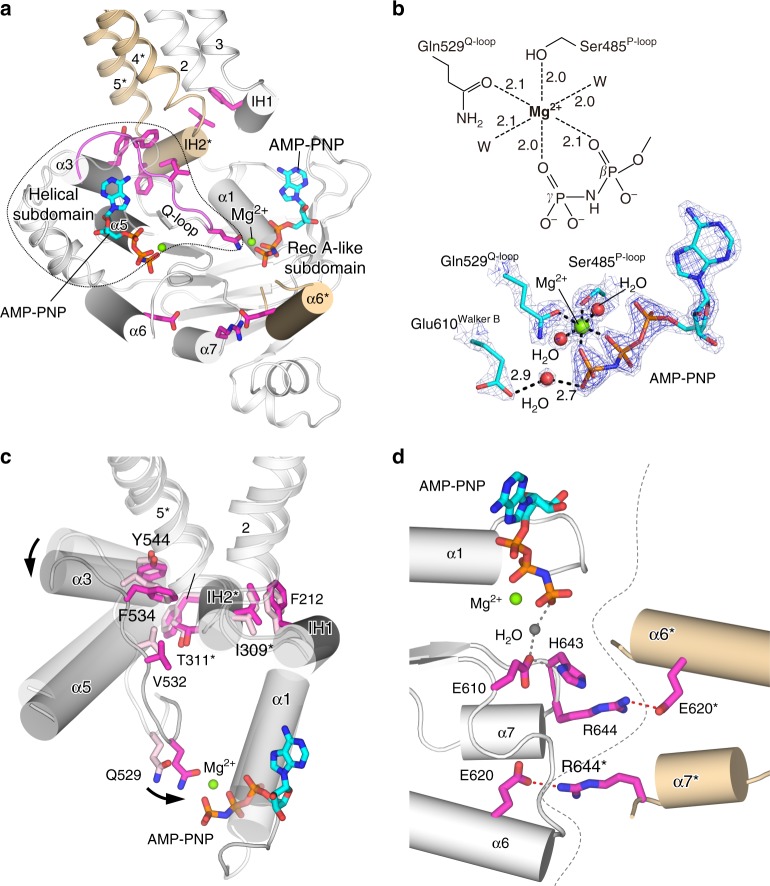
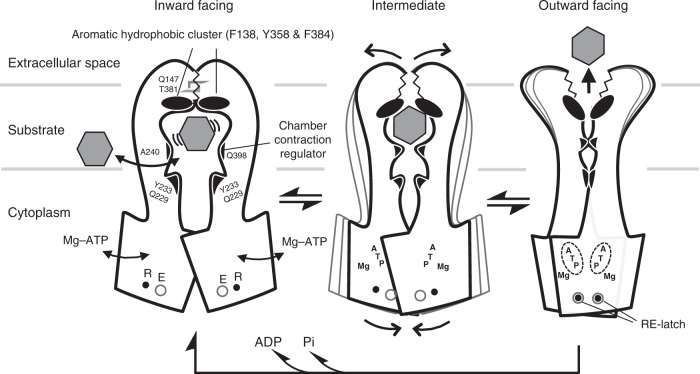
P-glycoprotein extrudes a large variety of xenobiotics from the cell, thereby protecting tissues from their toxic effects. The machinery underlying unidirectional multidrug pumping remains unknown, largely due to the lack of high-resolution structural information regarding the alternate conformational states of the molecule. Here we report a pair of structures of homodimeric P-glycoprotein: an outward-facing conformational state with bound nucleotide and an inward-facing apo state, at resolutions of 1.9 Å and 3.0 Å, respectively. Features that can be clearly visualized at this high resolution include ATP binding with octahedral coordination of Mg2+; an inner chamber that significantly changes in volume with the aid of tight connections among transmembrane helices (TM) 1, 3, and 6; a glutamate-arginine interaction that stabilizes the outward-facing conformation; and extensive interactions between TM1 and TM3, a property that distinguishes multidrug transporters from floppases. These structural elements are proposed to participate in the mechanism of the transporter.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
