

Recruitment of Vps34 PI3K and enrichment of PI3P phosphoinositide in the viral replication compartment is crucial for replication of a positive-strand RNA virus
- PMID: 30625229
- PMCID: PMC6342326
- DOI: 10.1371/journal.ppat.1007530
Recruitment of Vps34 PI3K and enrichment of PI3P phosphoinositide in the viral replication compartment is crucial for replication of a positive-strand RNA virus
Abstract
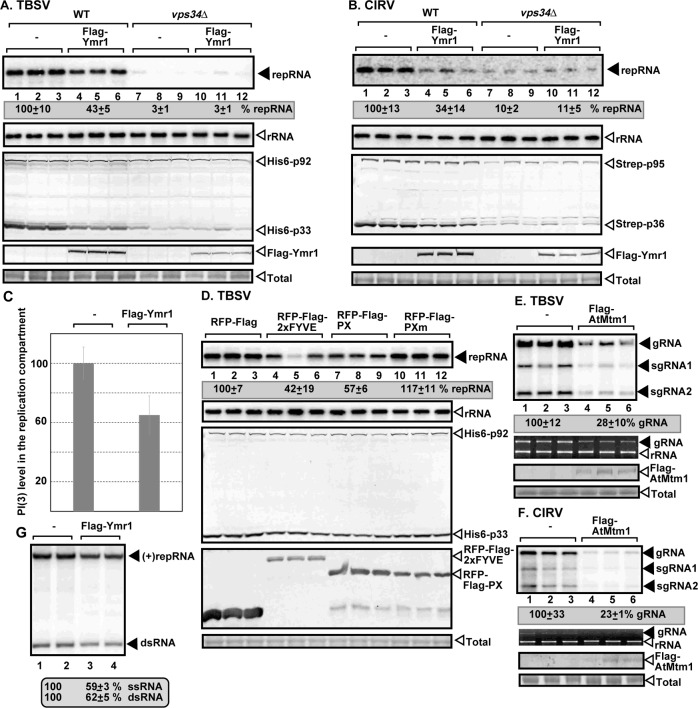
Tombusviruses depend on subversions of multiple host factors and retarget cellular pathways to support viral replication. In this work, we demonstrate that tomato bushy stunt virus (TBSV) and the closely-related carnation Italian ringspot virus (CIRV) recruit the cellular Vps34 phosphatidylinositol 3-kinase (PI3K) into the large viral replication compartment. The kinase function of Vps34 is critical for TBSV replication, suggesting that PI(3)P phosphoinositide is utilized by TBSV for building of the replication compartment. We also observed increased expression of Vps34 and the higher abundance of PI(3)P in the presence of the tombusviral replication proteins, which likely leads to more efficient tombusvirus replication. Accordingly, overexpression of PI(3)P phosphatase in yeast or plants inhibited TBSV replication on the peroxisomal membranes and CIRV replication on the mitochondrial membranes. Moreover, the purified PI(3)P phosphatase reduced TBSV replicase assembly in a cell-free system. Detection of PI(3)P with antibody or a bioprobe revealed the enrichment of PI(3)P in the replication compartment. Vps34 is directly recruited into the replication compartment through interaction with p33 replication protein. Gene deletion analysis in surrogate yeast host unraveled that TBSV replication requires the vesicle transport function of Vps34. In the absence of Vps34, TBSV cannot efficiently recruit the Rab5-positive early endosomes, which provide PE-rich membranes for membrane biogenesis of the TBSV replication compartment. We found that Vps34 and PI(3)P needed for the stability of the p33 replication protein, which is degraded by the 26S proteasome when PI(3)P abundance was decreased by an inhibitor of Vps34. In summary, Vps34 and PI(3)P are needed for providing the optimal microenvironment for the replication of the peroxisomal TBSV and the mitochondrial CIRV.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures










Similar articles
-
The retromer is co-opted to deliver lipid enzymes for the biogenesis of lipid-enriched tombusviral replication organelles.Proc Natl Acad Sci U S A. 2021 Jan 5;118(1):e2016066118. doi: 10.1073/pnas.2016066118. Proc Natl Acad Sci U S A. 2021. PMID: 33376201 Free PMC article.
-
Co-opted Cellular Sac1 Lipid Phosphatase and PI(4)P Phosphoinositide Are Key Host Factors during the Biogenesis of the Tombusvirus Replication Compartment.J Virol. 2020 Jun 1;94(12):e01979-19. doi: 10.1128/JVI.01979-19. Print 2020 Jun 1. J Virol. 2020. PMID: 32269127 Free PMC article.
-
Assembly-hub function of ER-localized SNARE proteins in biogenesis of tombusvirus replication compartment.PLoS Pathog. 2018 May 10;14(5):e1007028. doi: 10.1371/journal.ppat.1007028. eCollection 2018 May. PLoS Pathog. 2018. PMID: 29746582 Free PMC article.
-
Tombusvirus-Host Interactions: Co-Opted Evolutionarily Conserved Host Factors Take Center Court.Annu Rev Virol. 2016 Sep 29;3(1):491-515. doi: 10.1146/annurev-virology-110615-042312. Epub 2016 Aug 8. Annu Rev Virol. 2016. PMID: 27578441 Review.
-
Tombusvirus polymerase: Structure and function.Virus Res. 2017 Apr 15;234:74-86. doi: 10.1016/j.virusres.2017.01.012. Epub 2017 Jan 19. Virus Res. 2017. PMID: 28111194 Review.
Cited by
-
Dynamic interplay between the co-opted Fis1 mitochondrial fission protein and membrane contact site proteins in supporting tombusvirus replication.PLoS Pathog. 2021 Mar 16;17(3):e1009423. doi: 10.1371/journal.ppat.1009423. eCollection 2021 Mar. PLoS Pathog. 2021. PMID: 33725015 Free PMC article.
-
Host proviral and antiviral factors for SARS-CoV-2.Virus Genes. 2021 Dec;57(6):475-488. doi: 10.1007/s11262-021-01869-2. Epub 2021 Sep 11. Virus Genes. 2021. PMID: 34510331 Free PMC article. Review.
-
Subversion of selective autophagy for the biogenesis of tombusvirus replication organelles inhibits autophagy.PLoS Pathog. 2024 Mar 14;20(3):e1012085. doi: 10.1371/journal.ppat.1012085. eCollection 2024 Mar. PLoS Pathog. 2024. PMID: 38484009 Free PMC article.
-
The retromer is co-opted to deliver lipid enzymes for the biogenesis of lipid-enriched tombusviral replication organelles.Proc Natl Acad Sci U S A. 2021 Jan 5;118(1):e2016066118. doi: 10.1073/pnas.2016066118. Proc Natl Acad Sci U S A. 2021. PMID: 33376201 Free PMC article.
-
Mobilization of nuclear antiviral factors by exportin XPO1 via the actin network inhibits RNA virus replication.PLoS Pathog. 2025 Aug 19;21(8):e1012841. doi: 10.1371/journal.ppat.1012841. eCollection 2025 Aug. PLoS Pathog. 2025. PMID: 40828816 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
