

LHX2- and LDB1-mediated trans interactions regulate olfactory receptor choice
- PMID: 30626972
- PMCID: PMC6436840
- DOI: 10.1038/s41586-018-0845-0
LHX2- and LDB1-mediated trans interactions regulate olfactory receptor choice
Abstract
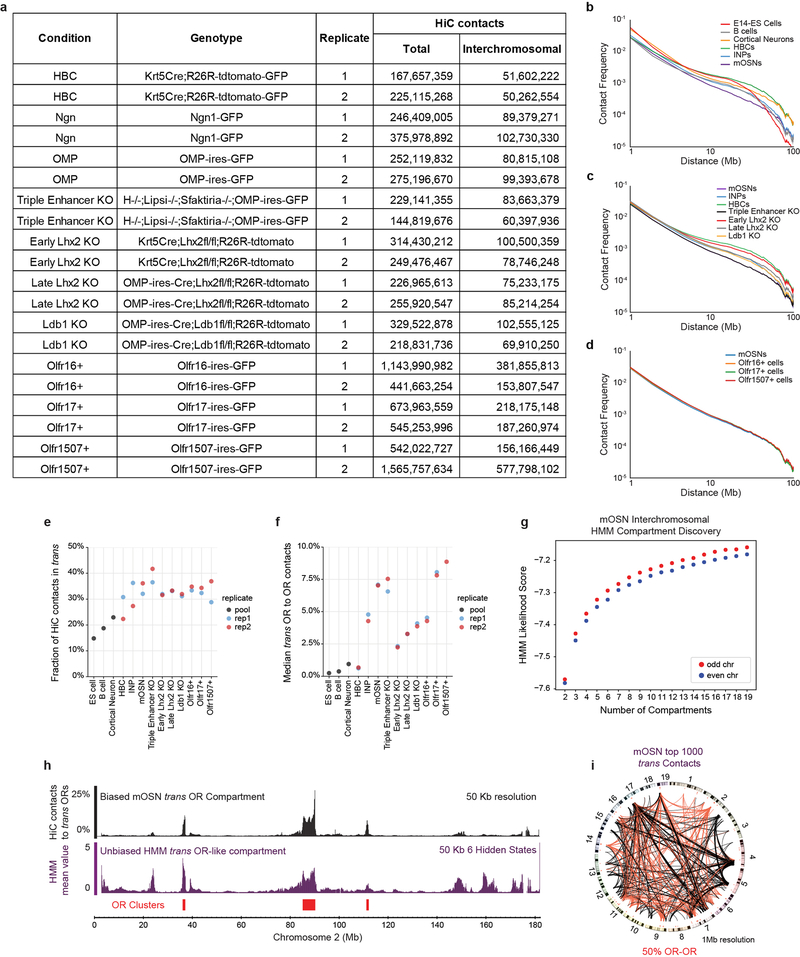
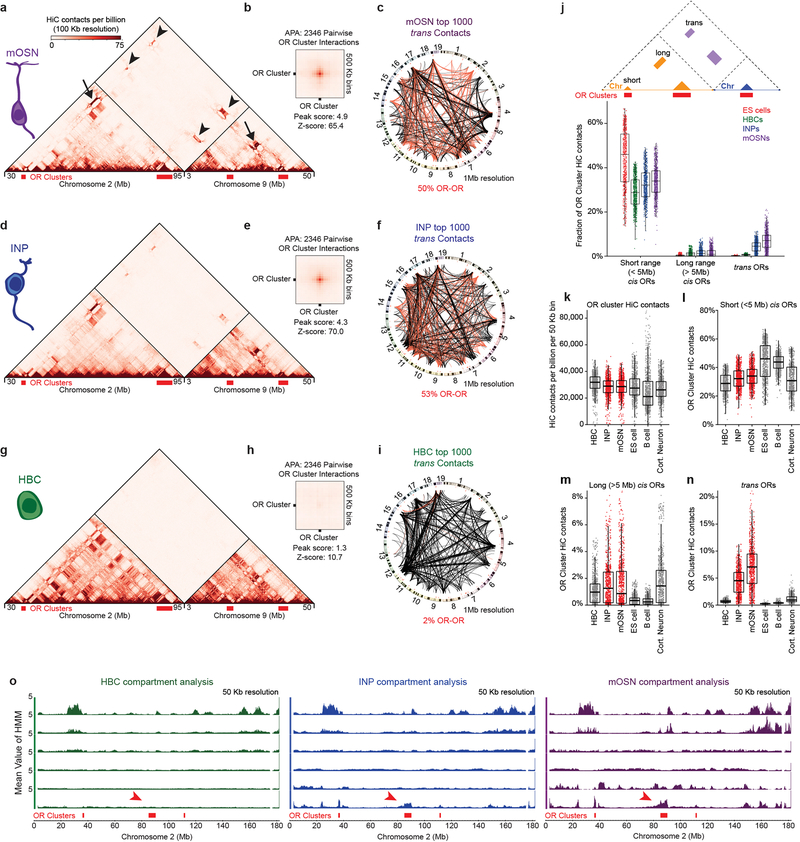
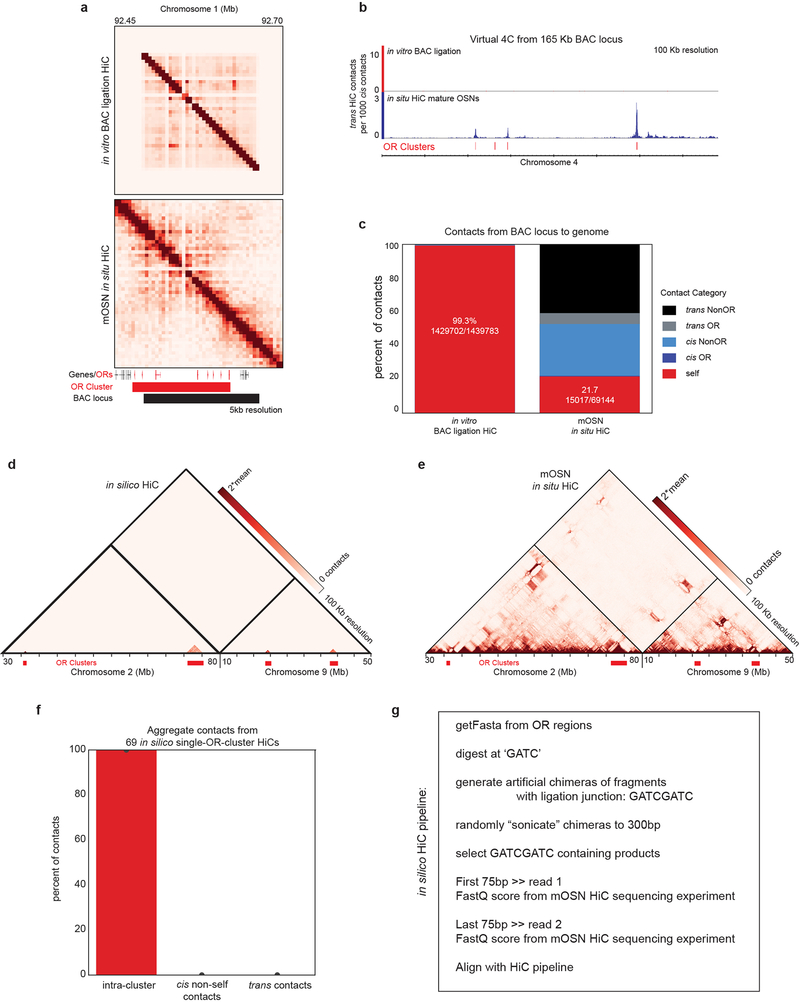
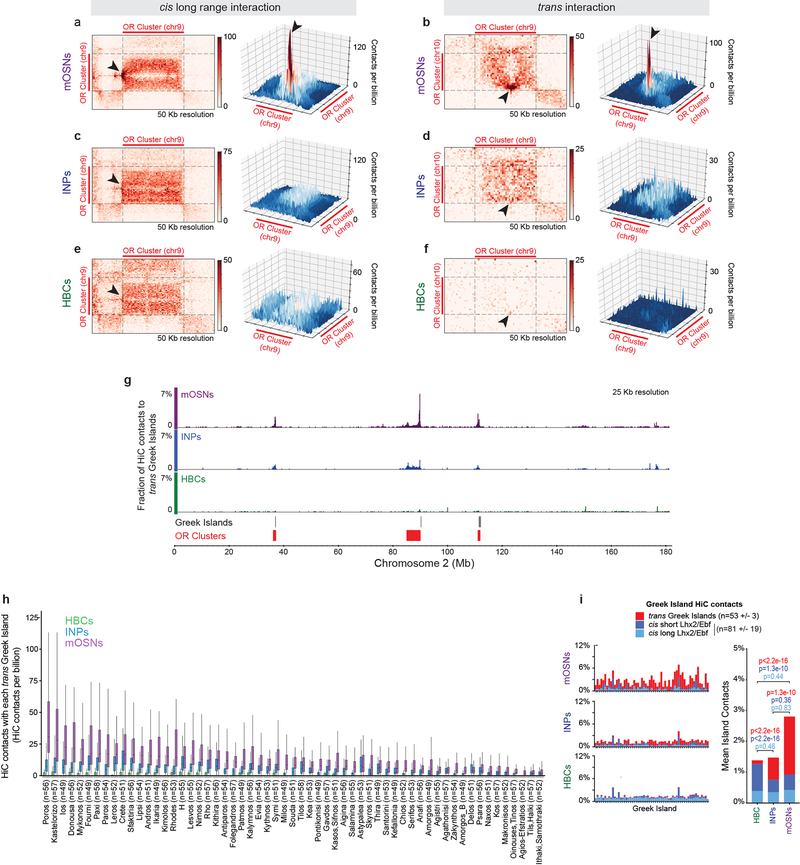
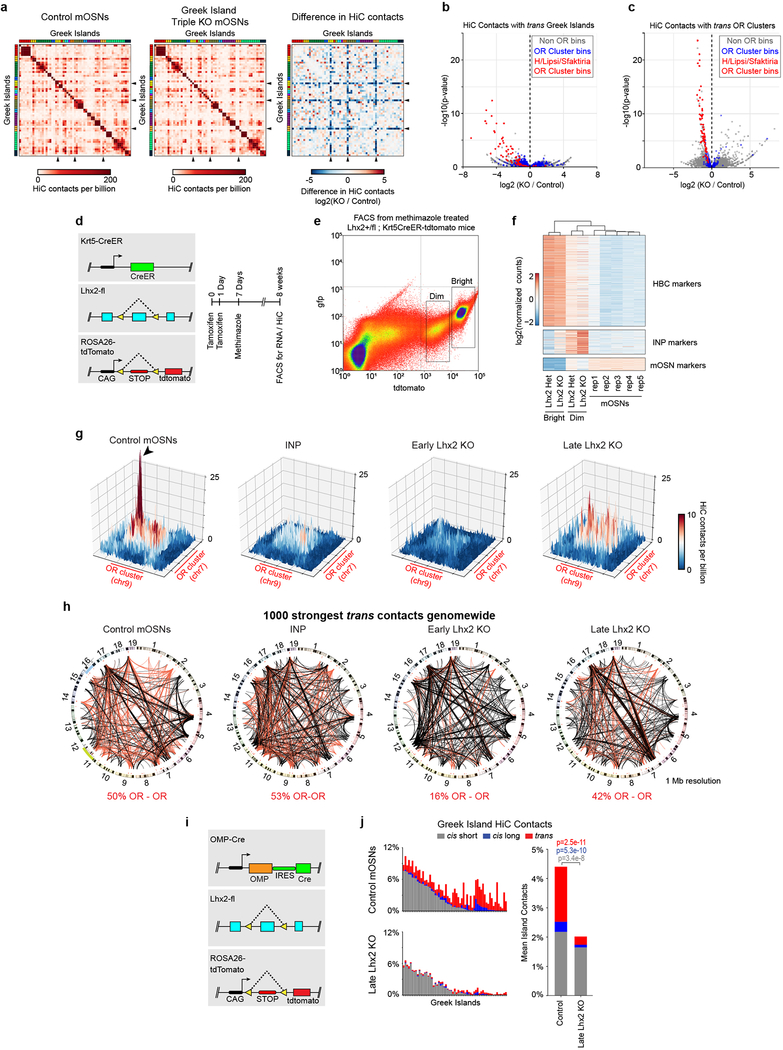
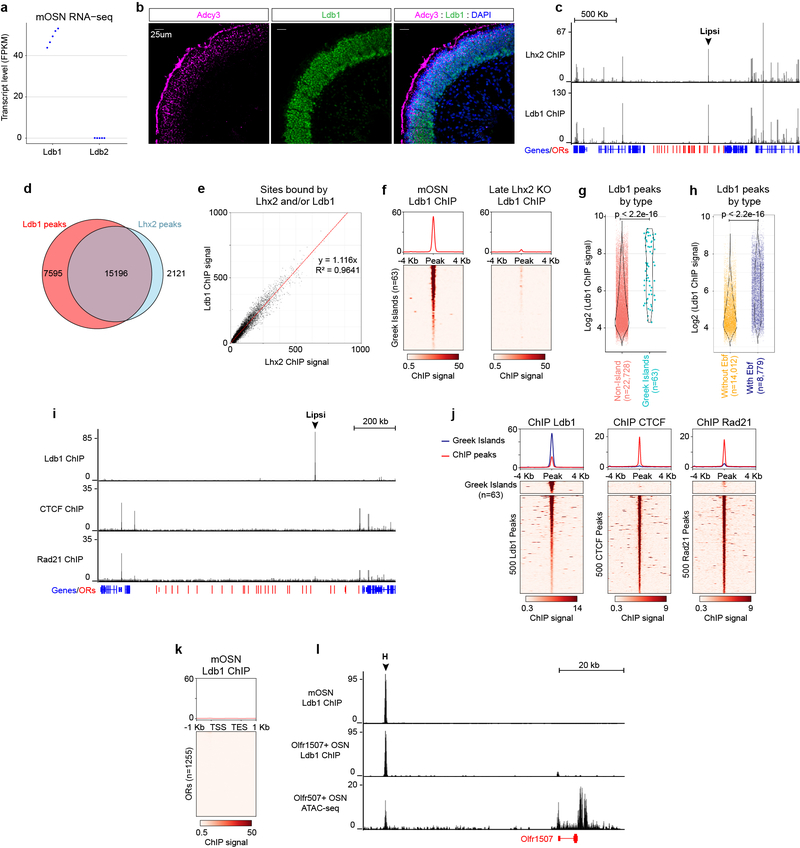
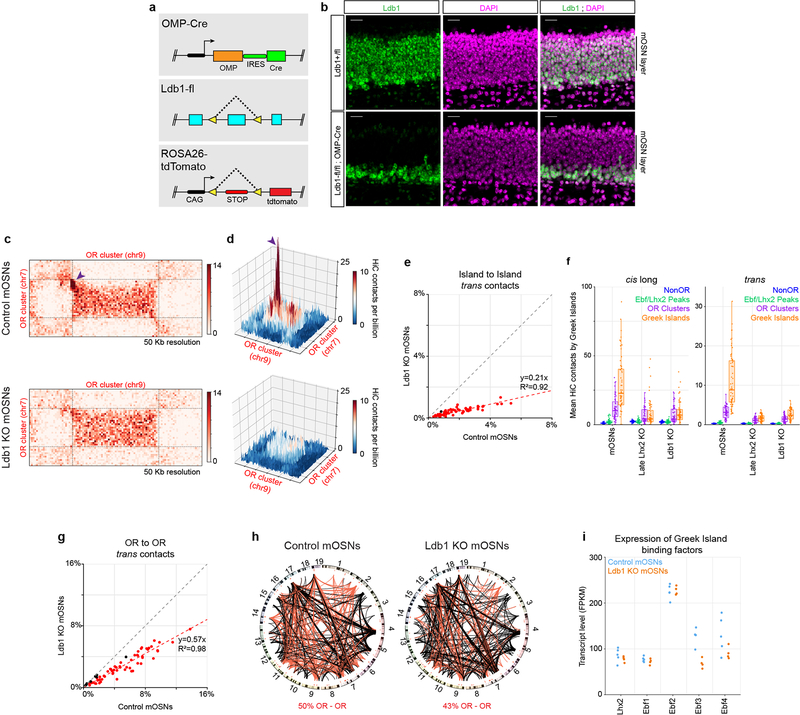
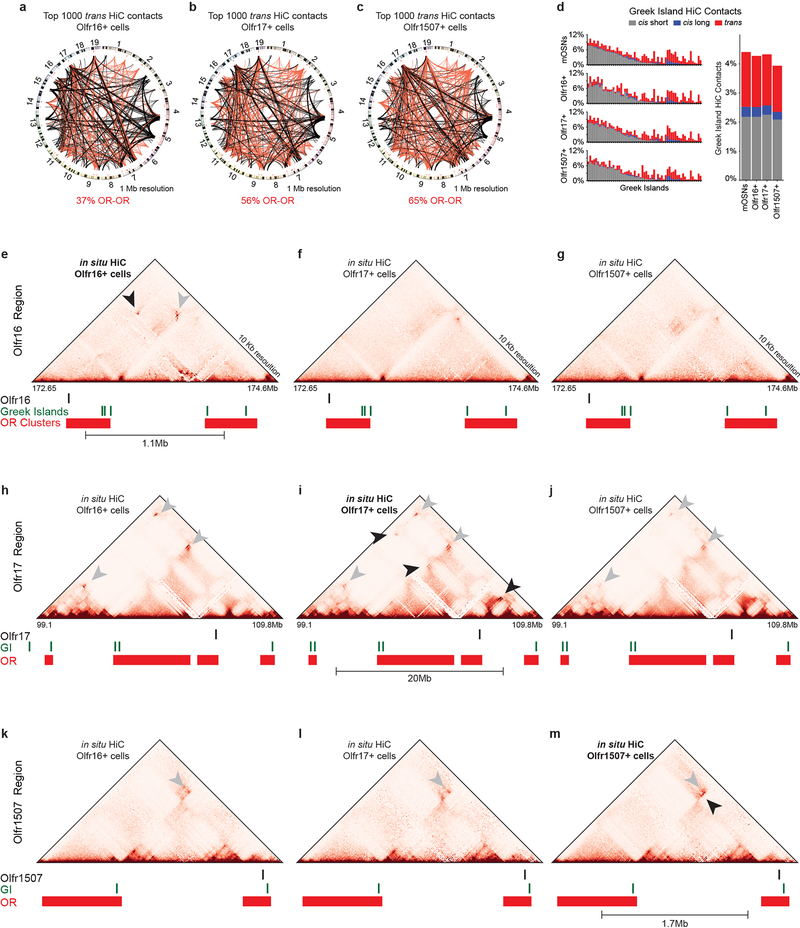
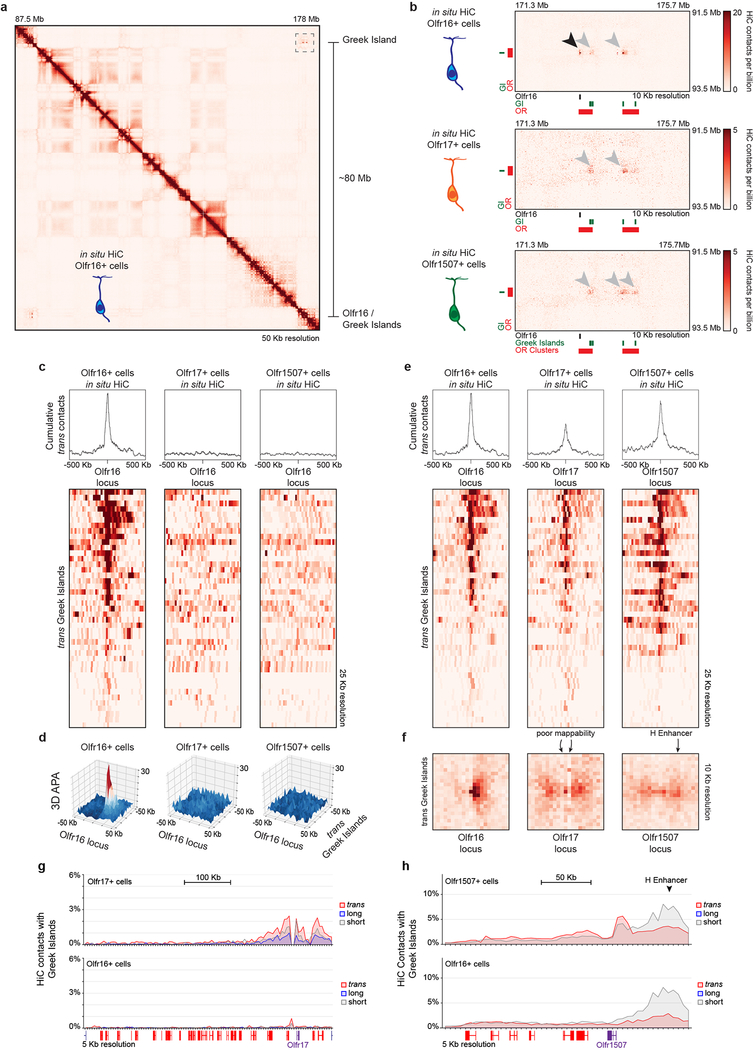
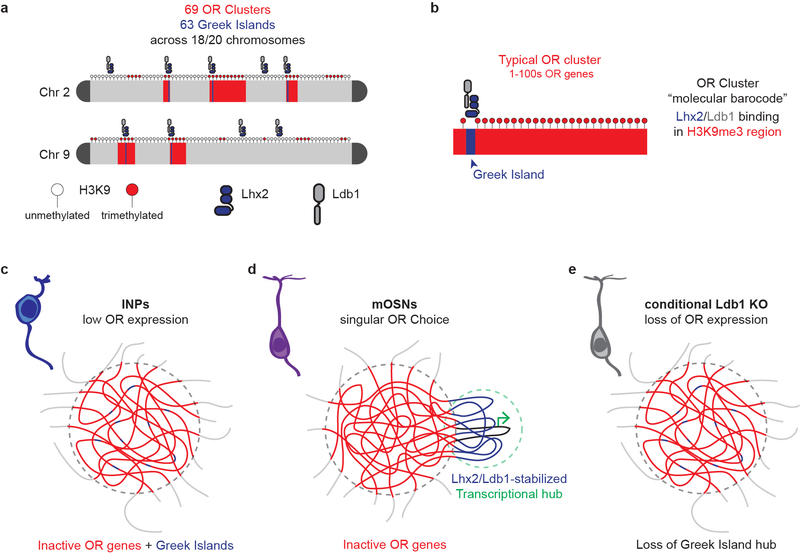
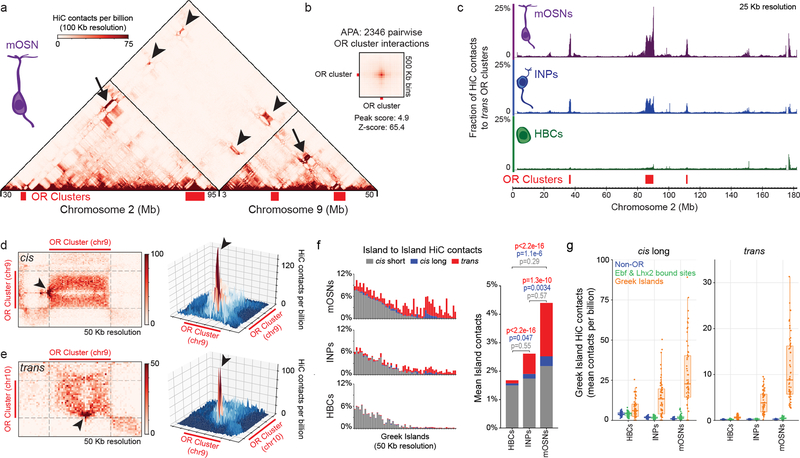
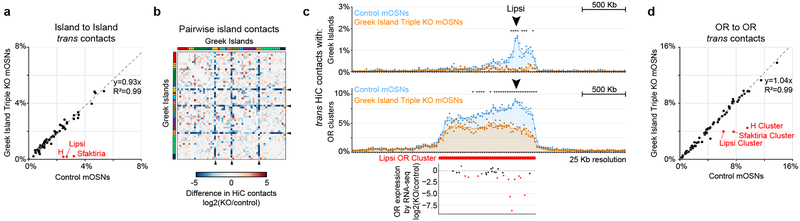
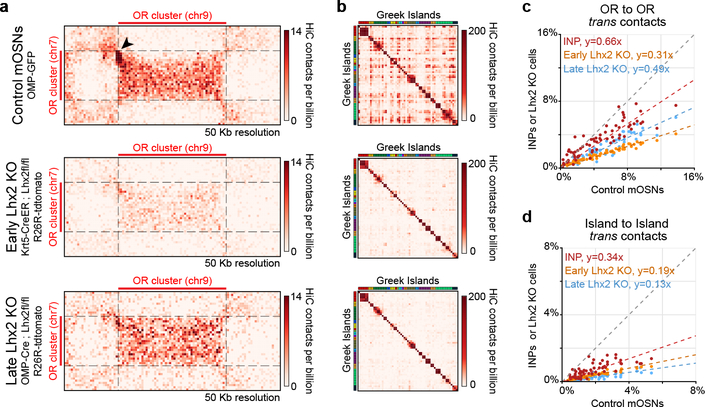
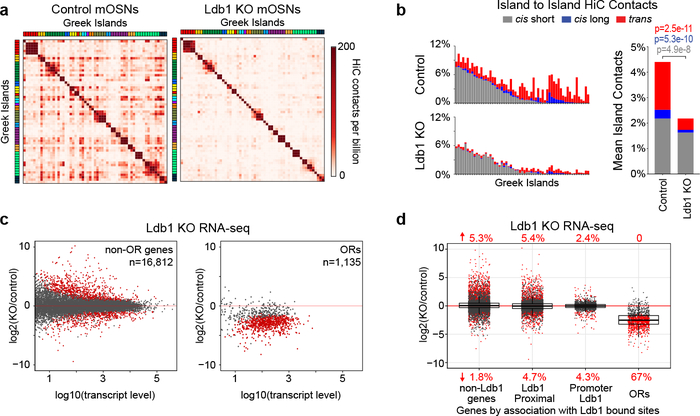
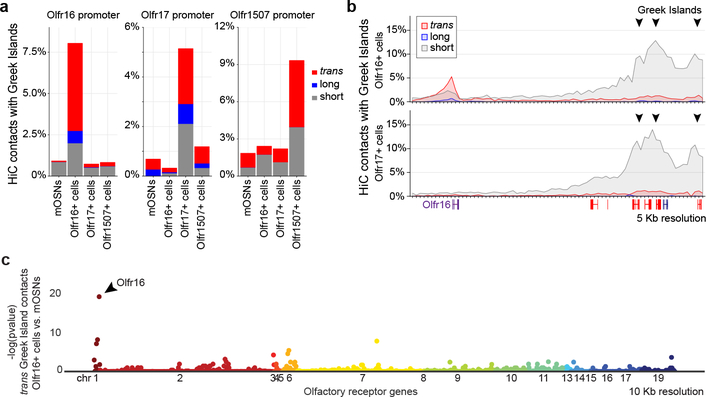
The genome is partitioned into topologically associated domains and genomic compartments with shared chromatin valence. This architecture is constrained by the DNA polymer, which precludes interactions between genes on different chromosomes. Here we report a marked divergence from this pattern of nuclear organization that occurs in mouse olfactory sensory neurons. Chromatin conformation capture using in situ Hi-C on fluorescence-activated cell-sorted olfactory sensory neurons and their progenitors shows that olfactory receptor gene clusters from 18 chromosomes make specific and robust interchromosomal contacts that increase with differentiation of the cells. These contacts are orchestrated by intergenic olfactory receptor enhancers, the 'Greek islands', which first contribute to the formation of olfactory receptor compartments and then form a multi-chromosomal super-enhancer that associates with the single active olfactory receptor gene. The Greek-island-bound transcription factor LHX2 and adaptor protein LDB1 regulate the assembly and maintenance of olfactory receptor compartments, Greek island hubs and olfactory receptor transcription, providing mechanistic insights into and functional support for the role of trans interactions in gene expression.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
Comment in
-
Chromosomes come together to help mice distinguish odours.Nature. 2019 Jan;565(7740):439-440. doi: 10.1038/d41586-019-00010-6. Nature. 2019. PMID: 30666054 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
