Neuropeptide and Small Transmitter Coexistence: Fundamental Studies and Relevance to Mental Illness
- PMID: 30627087
- PMCID: PMC6309708
- DOI: 10.3389/fncir.2018.00106
Neuropeptide and Small Transmitter Coexistence: Fundamental Studies and Relevance to Mental Illness
Abstract
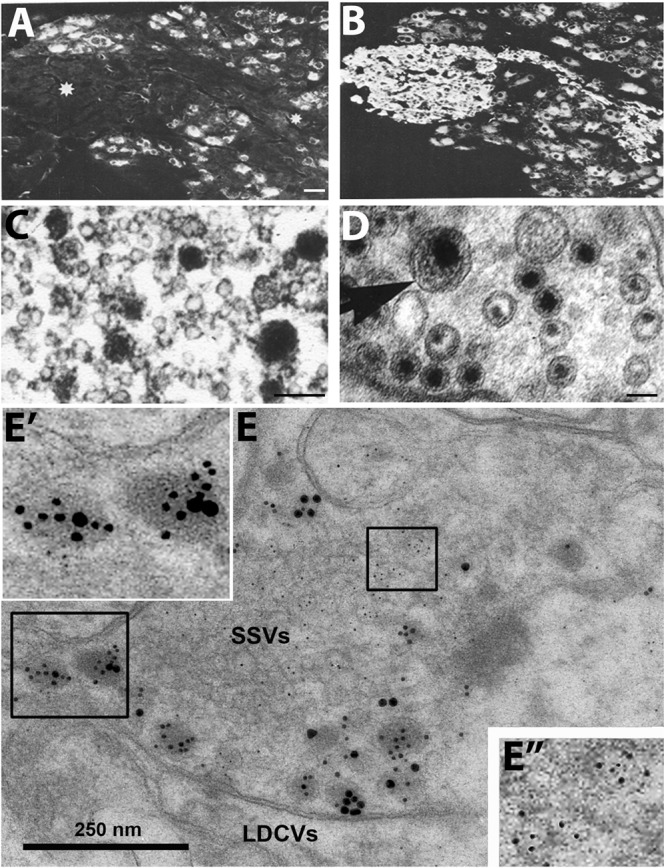
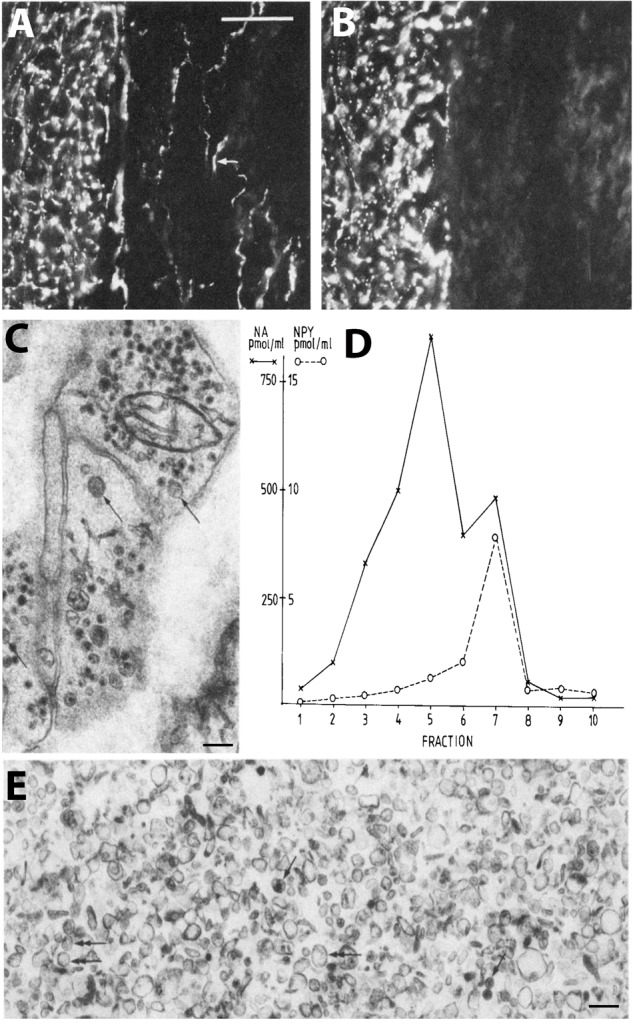
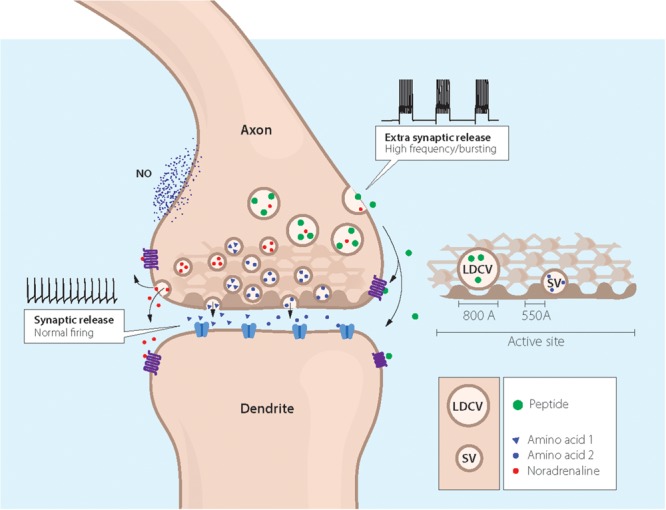
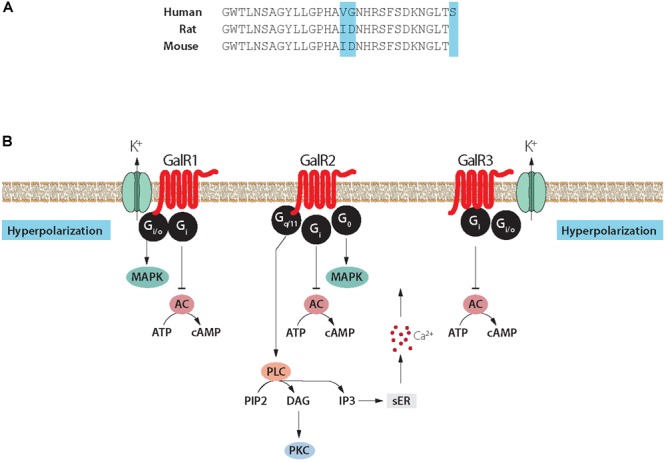
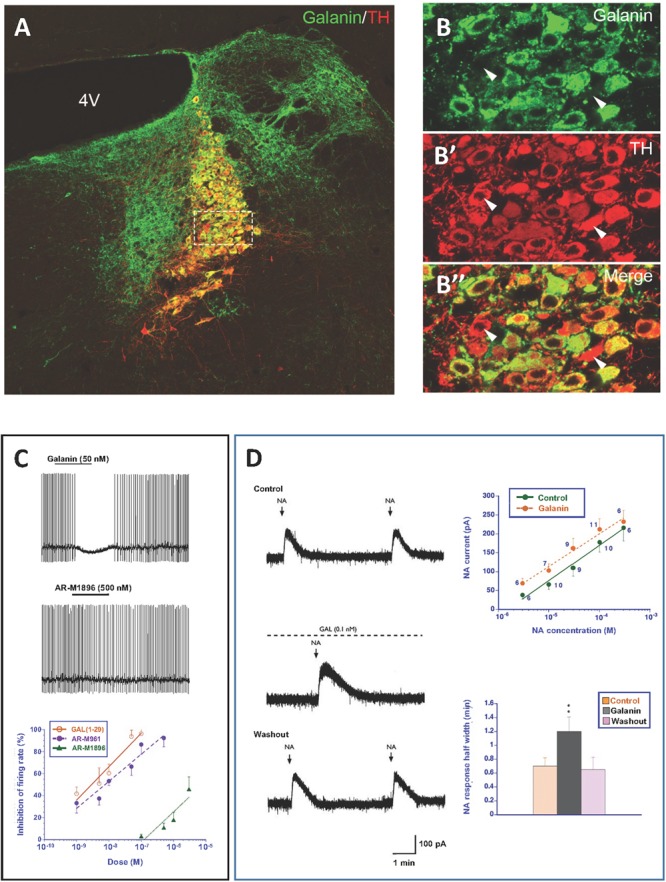
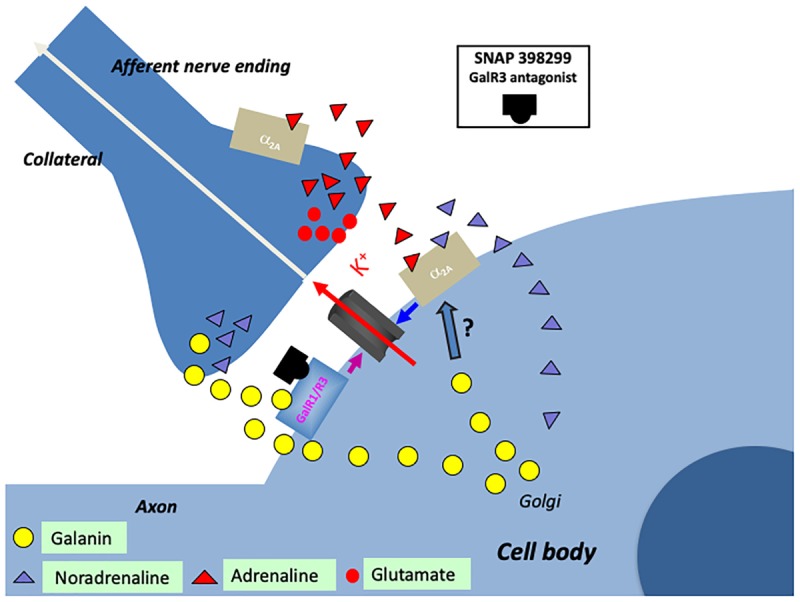
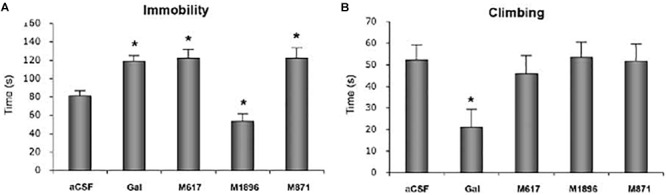
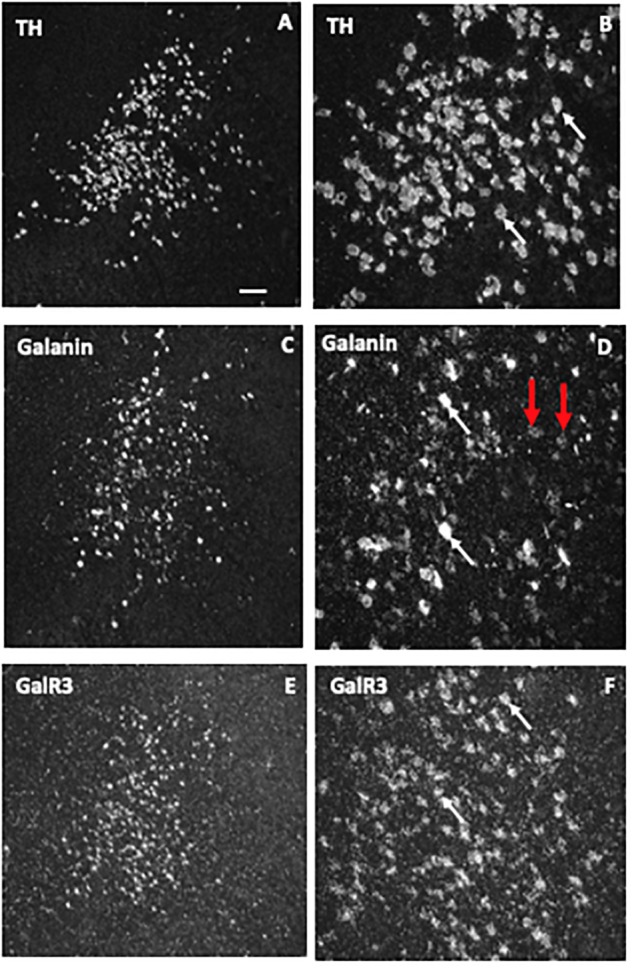
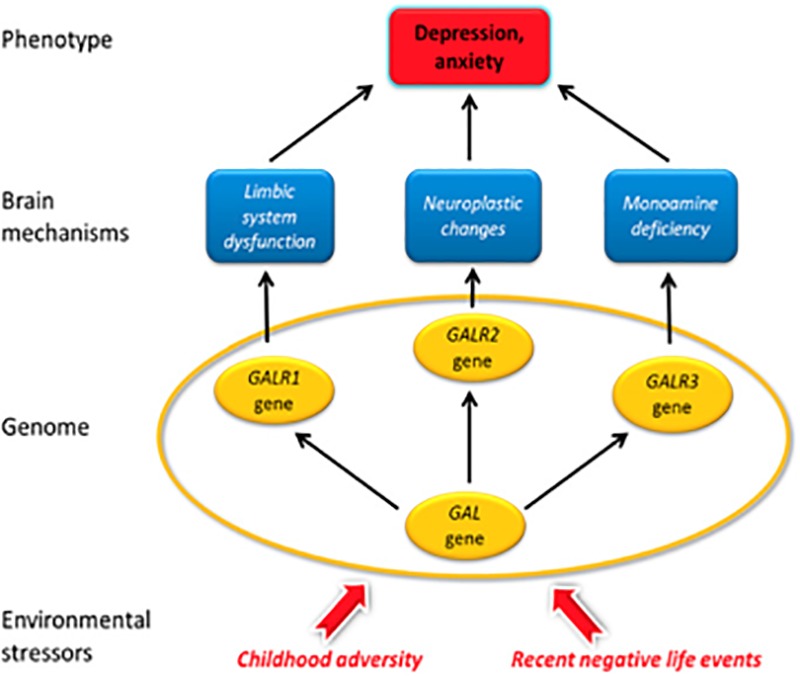
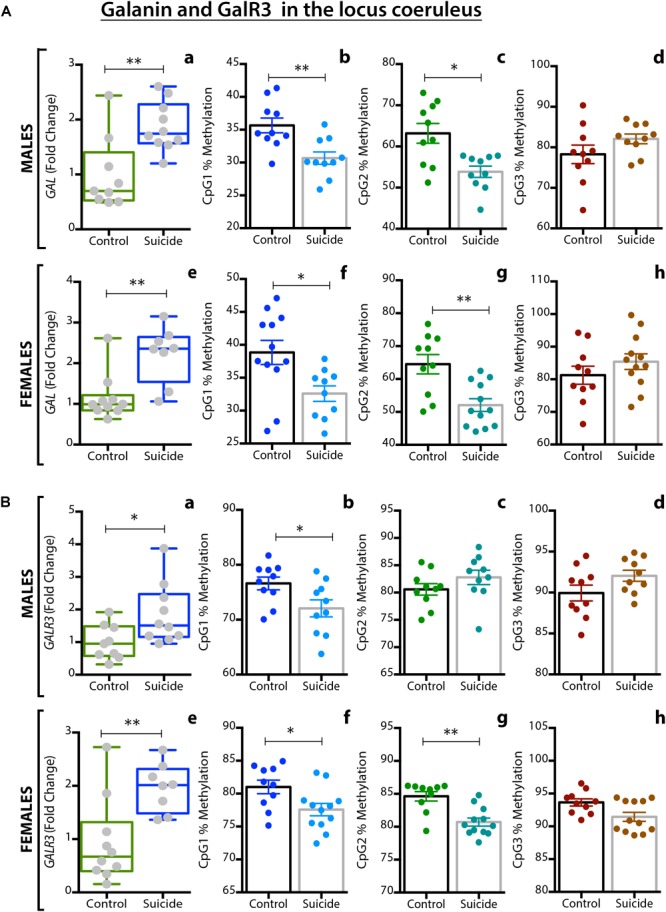
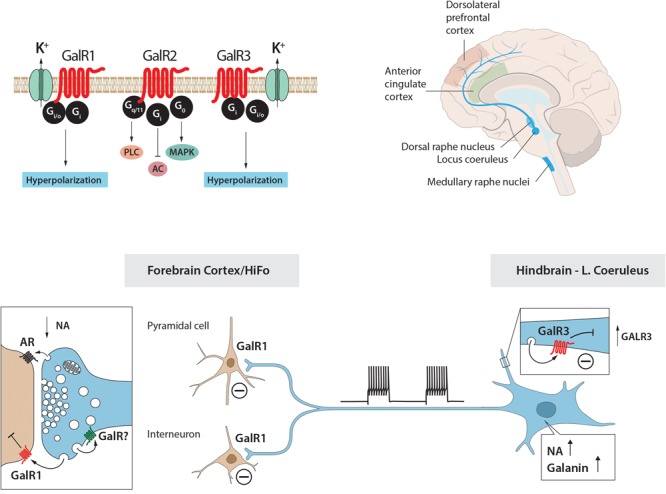
Neuropeptides are auxiliary messenger molecules that always co-exist in nerve cells with one or more small molecule (classic) neurotransmitters. Neuropeptides act both as transmitters and trophic factors, and play a role particularly when the nervous system is challenged, as by injury, pain or stress. Here neuropeptides and coexistence in mammals are reviewed, but with special focus on the 29/30 amino acid galanin and its three receptors GalR1, -R2 and -R3. In particular, galanin's role as a co-transmitter in both rodent and human noradrenergic locus coeruleus (LC) neurons is addressed. Extensive experimental animal data strongly suggest a role for the galanin system in depression-like behavior. The translational potential of these results was tested by studying the galanin system in postmortem human brains, first in normal brains, and then in a comparison of five regions of brains obtained from depressed people who committed suicide, and from matched controls. The distribution of galanin and the four galanin system transcripts in the normal human brain was determined, and selective and parallel changes in levels of transcripts and DNA methylation for galanin and its three receptors were assessed in depressed patients who committed suicide: upregulation of transcripts, e.g., for galanin and GalR3 in LC, paralleled by a decrease in DNA methylation, suggesting involvement of epigenetic mechanisms. It is hypothesized that, when exposed to severe stress, the noradrenergic LC neurons fire in bursts and release galanin from their soma/dendrites. Galanin then acts on somato-dendritic, inhibitory galanin autoreceptors, opening potassium channels and inhibiting firing. The purpose of these autoreceptors is to act as a 'brake' to prevent overexcitation, a brake that is also part of resilience to stress that protects against depression. Depression then arises when the inhibition is too strong and long lasting - a maladaption, allostatic load, leading to depletion of NA levels in the forebrain. It is suggested that disinhibition by a galanin antagonist may have antidepressant activity by restoring forebrain NA levels. A role of galanin in depression is also supported by a recent candidate gene study, showing that variants in genes for galanin and its three receptors confer increased risk of depression and anxiety in people who experienced childhood adversity or recent negative life events. In summary, galanin, a neuropeptide coexisting in LC neurons, may participate in the mechanism underlying resilience against a serious and common disorder, MDD. Existing and further results may lead to an increased understanding of how this illness develops, which in turn could provide a basis for its treatment.
Keywords: allostatic load; epigenetics; galanin; locus coeruleus; major depression disorder; neuropeptides; resilience.
Figures