

Centromere and Pericentromere Transcription: Roles and Regulation … in Sickness and in Health
- PMID: 30627137
- PMCID: PMC6309819
- DOI: 10.3389/fgene.2018.00674
Centromere and Pericentromere Transcription: Roles and Regulation … in Sickness and in Health
Abstract
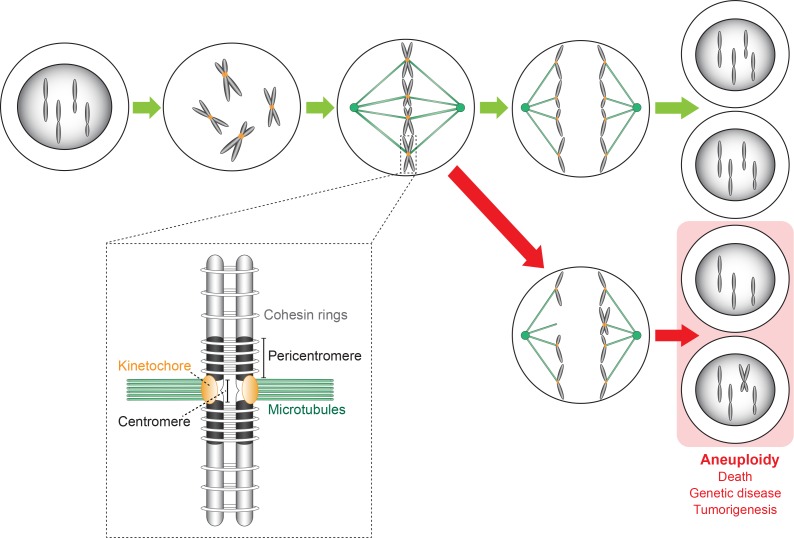
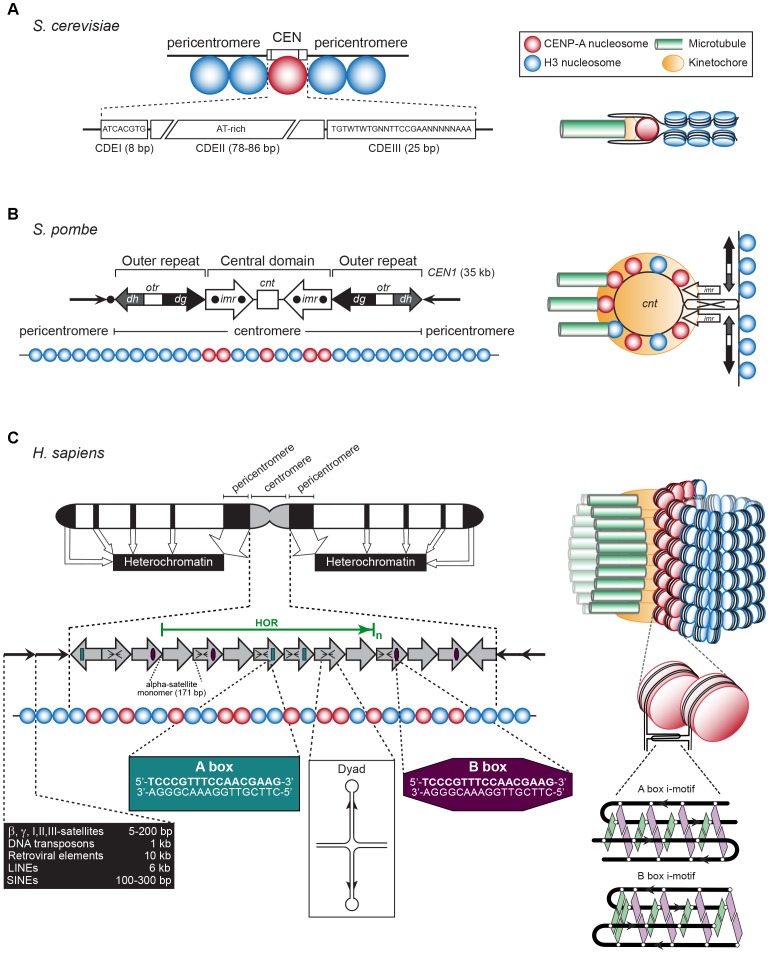
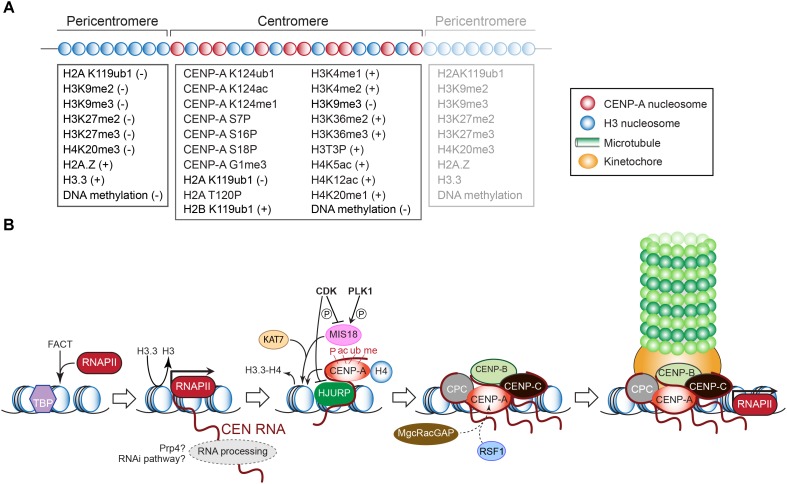
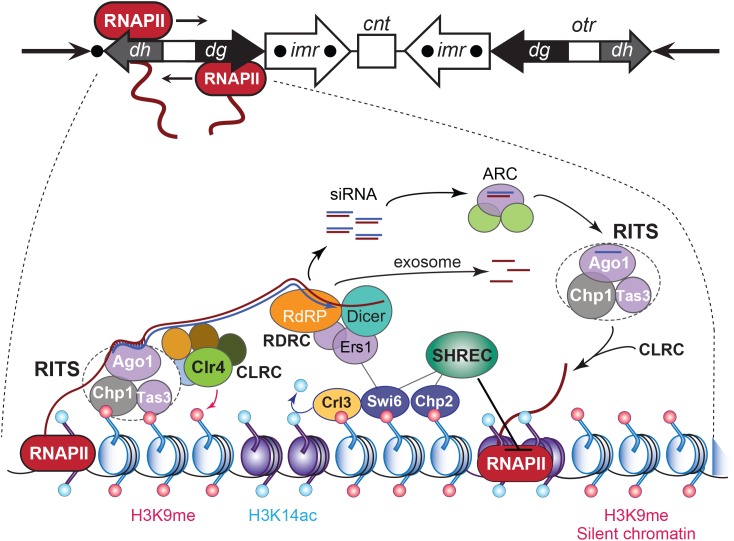
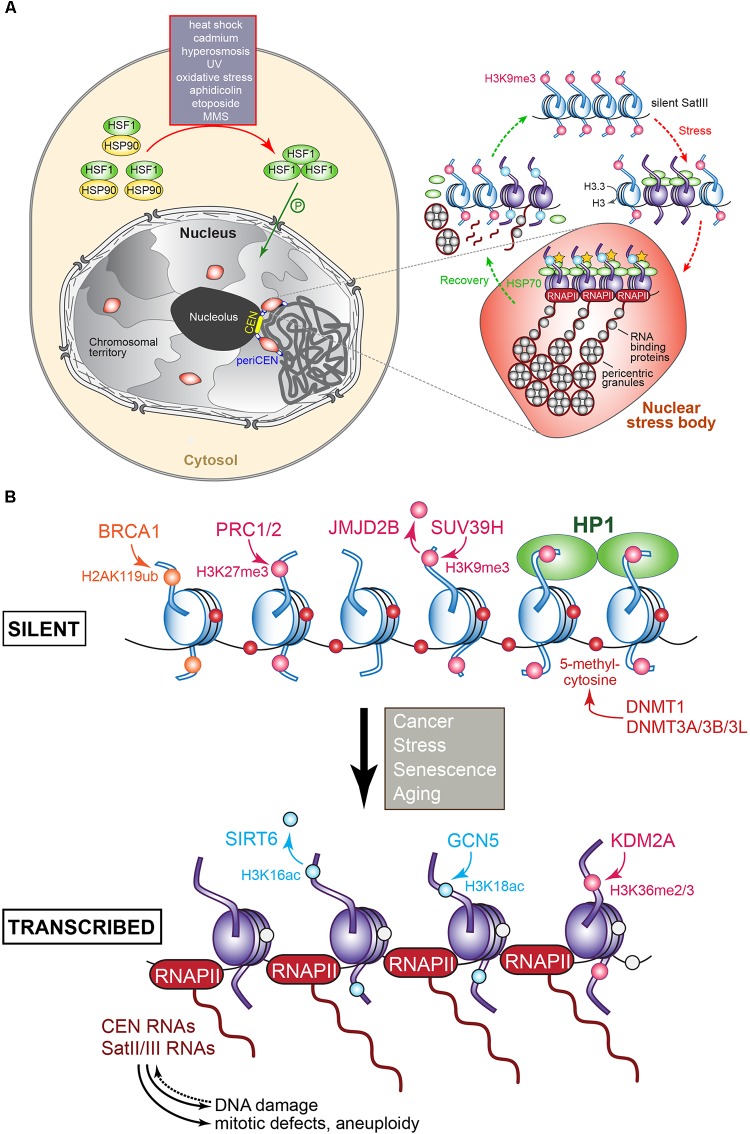
The chromosomal loci known as centromeres (CEN) mediate the equal distribution of the duplicated genome between both daughter cells. Specifically, centromeres recruit a protein complex named the kinetochore, that bi-orients the replicated chromosome pairs to the mitotic or meiotic spindle structure. The paired chromosomes are then separated, and the individual chromosomes segregate in opposite direction along the regressing spindle into each daughter cell. Erroneous kinetochore assembly or activity produces aneuploid cells that contain an abnormal number of chromosomes. Aneuploidy may incite cell death, developmental defects (including genetic syndromes), and cancer (>90% of all cancer cells are aneuploid). While kinetochores and their activities have been preserved through evolution, the CEN DNA sequences have not. Hence, to be recognized as sites for kinetochore assembly, CEN display conserved structural themes. In addition, CEN nucleosomes enclose a CEN-exclusive variant of histone H3, named CENP-A, and carry distinct epigenetic labels on CENP-A and the other CEN histone proteins. Through the cell cycle, CEN are transcribed into non-coding RNAs. After subsequent processing, they become key components of the CEN chromatin by marking the CEN locus and by stably anchoring the CEN-binding kinetochore proteins. CEN transcription is tightly regulated, of low intensity, and essential for differentiation and development. Under- or overexpression of CEN transcripts, as documented for myriad cancers, provoke chromosome missegregation and aneuploidy. CEN are genetically stable and fully competent only when they are insulated from the surrounding, pericentromeric chromatin, which must be silenced. We will review CEN transcription and its contribution to faithful kinetochore function. We will further discuss how pericentromeric chromatin is silenced by RNA processing and transcriptionally repressive chromatin marks. We will report on the transcriptional misregulation of (peri)centromeres during stress, natural aging, and disease and reflect on whether their transcripts can serve as future diagnostic tools and anti-cancer targets in the clinic.
Keywords: centromere; heterochromatin; kinetochore; long non-coding RNA; pericentromere; transcription.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources