

Plastid Genomes and Proteins Illuminate the Evolution of Eustigmatophyte Algae and Their Bacterial Endosymbionts
- PMID: 30629162
- PMCID: PMC6367104
- DOI: 10.1093/gbe/evz004
Plastid Genomes and Proteins Illuminate the Evolution of Eustigmatophyte Algae and Their Bacterial Endosymbionts
Abstract
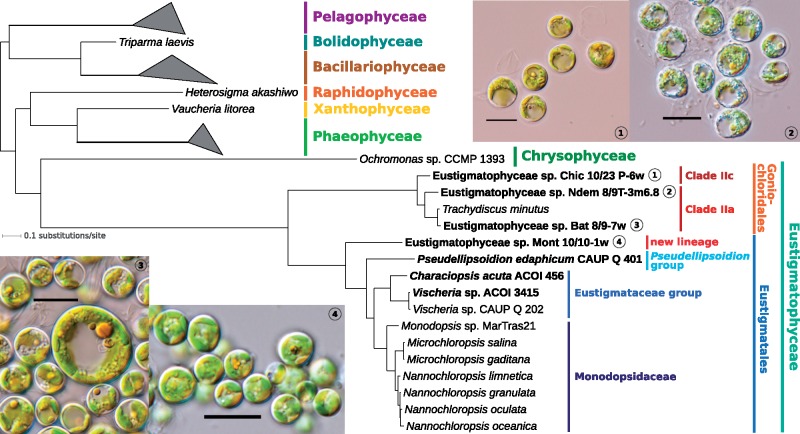
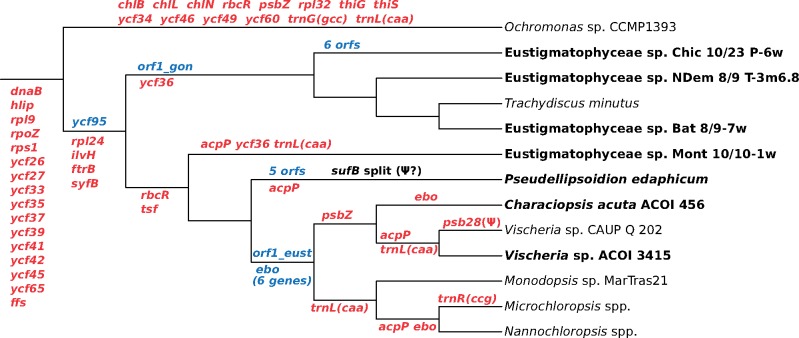
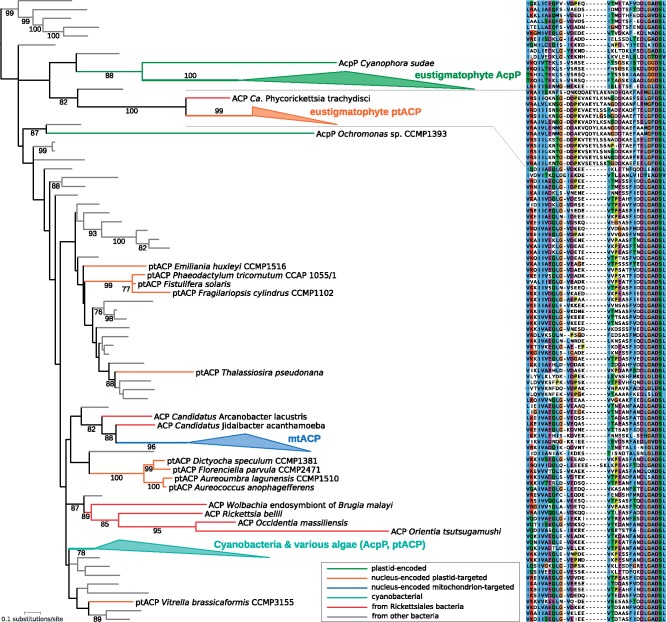
Eustigmatophytes, a class of stramenopile algae (ochrophytes), include not only the extensively studied biotechnologically important genus Nannochloropsis but also a rapidly expanding diversity of lineages with much less well characterized biology. Recent discoveries have led to exciting additions to our knowledge about eustigmatophytes. Some proved to harbor bacterial endosymbionts representing a novel genus, Candidatus Phycorickettsia, and an operon of unclear function (ebo) obtained by horizontal gene transfer from the endosymbiont lineage was found in the plastid genomes of still other eustigmatophytes. To shed more light on the latter event, as well as to generally improve our understanding of the eustigmatophyte evolutionary history, we sequenced plastid genomes of seven phylogenetically diverse representatives (including new isolates representing undescribed taxa). A phylogenomic analysis of plastid genome-encoded proteins resolved the phylogenetic relationships among the main eustigmatophyte lineages and provided a framework for the interpretation of plastid gene gains and losses in the group. The ebo operon gain was inferred to have probably occurred within the order Eustigmatales, after the divergence of the two basalmost lineages (a newly discovered hitherto undescribed strain and the Pseudellipsoidion group). When looking for nuclear genes potentially compensating for plastid gene losses, we noticed a gene for a plastid-targeted acyl carrier protein that was apparently acquired by horizontal gene transfer from Phycorickettsia. The presence of this gene in all eustigmatophytes studied, including representatives of both principal clades (Eustigmatales and Goniochloridales), is a genetic footprint indicating that the eustigmatophyte-Phycorickettsia partnership started no later than in the last eustigmatophyte common ancestor.
Figures


Similar articles
-
A gene transfer event suggests a long-term partnership between eustigmatophyte algae and a novel lineage of endosymbiotic bacteria.ISME J. 2018 Sep;12(9):2163-2175. doi: 10.1038/s41396-018-0177-y. Epub 2018 Jun 7. ISME J. 2018. PMID: 29880910 Free PMC article.
-
Redefining Chlorobotryaceae as one of the principal and most diverse lineages of eustigmatophyte algae.Mol Phylogenet Evol. 2022 Dec;177:107607. doi: 10.1016/j.ympev.2022.107607. Epub 2022 Aug 11. Mol Phylogenet Evol. 2022. PMID: 35963589
-
Updating algal evolutionary relationships through plastid genome sequencing: did alveolate plastids emerge through endosymbiosis of an ochrophyte?Sci Rep. 2015 May 28;5:10134. doi: 10.1038/srep10134. Sci Rep. 2015. PMID: 26017773 Free PMC article.
-
A new scenario of plastid evolution: plastid primary endosymbiosis before the divergence of the "Plantae," emended.J Plant Res. 2005 Aug;118(4):247-55. doi: 10.1007/s10265-005-0219-1. Epub 2005 Jul 20. J Plant Res. 2005. PMID: 16032387 Review.
-
The origin and establishment of the plastid in algae and plants.Annu Rev Genet. 2007;41:147-68. doi: 10.1146/annurev.genet.41.110306.130134. Annu Rev Genet. 2007. PMID: 17600460 Review.
Cited by
-
Identification of Eukaryotic Microalgal Strains.J Appl Phycol. 2020 Oct;32(5):2699-2709. doi: 10.1007/s10811-020-02190-5. Epub 2020 Jul 15. J Appl Phycol. 2020. PMID: 33542589 Free PMC article.
-
Host Range and Coding Potential of Eukaryotic Giant Viruses.Viruses. 2020 Nov 21;12(11):1337. doi: 10.3390/v12111337. Viruses. 2020. PMID: 33233432 Free PMC article. Review.
-
Comparative Plastid Genomics of Non-Photosynthetic Chrysophytes: Genome Reduction and Compaction.Front Plant Sci. 2020 Sep 10;11:572703. doi: 10.3389/fpls.2020.572703. eCollection 2020. Front Plant Sci. 2020. PMID: 33013997 Free PMC article.
-
Implicating the red body of Nannochloropsis in forming the recalcitrant cell wall polymer algaenan.Nat Commun. 2024 Jun 27;15(1):5456. doi: 10.1038/s41467-024-49277-y. Nat Commun. 2024. PMID: 38937455 Free PMC article.
-
Organellar genome comparisons of Sargassum polycystum and S. plagiophyllum (Fucales, Phaeophyceae) with other Sargassum species.BMC Genomics. 2022 Sep 2;23(1):629. doi: 10.1186/s12864-022-08862-5. BMC Genomics. 2022. PMID: 36050627 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
