

Activated PMN Exosomes: Pathogenic Entities Causing Matrix Destruction and Disease in the Lung
- PMID: 30633902
- PMCID: PMC6368091
- DOI: 10.1016/j.cell.2018.12.002
Activated PMN Exosomes: Pathogenic Entities Causing Matrix Destruction and Disease in the Lung
Abstract
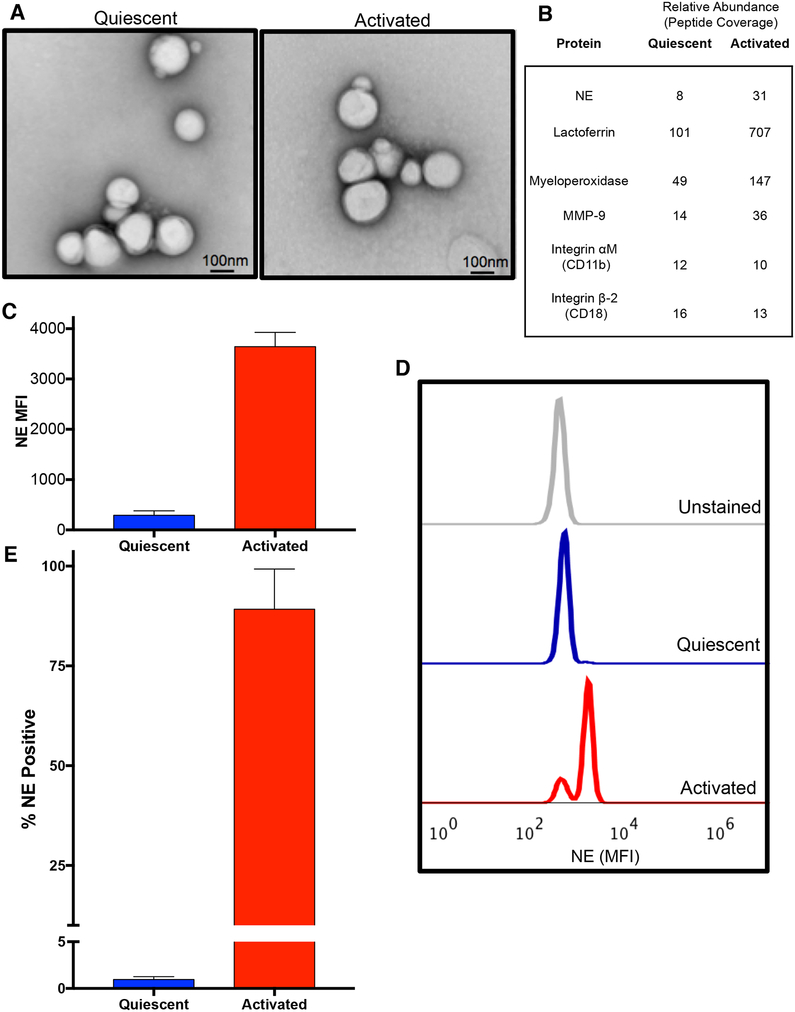
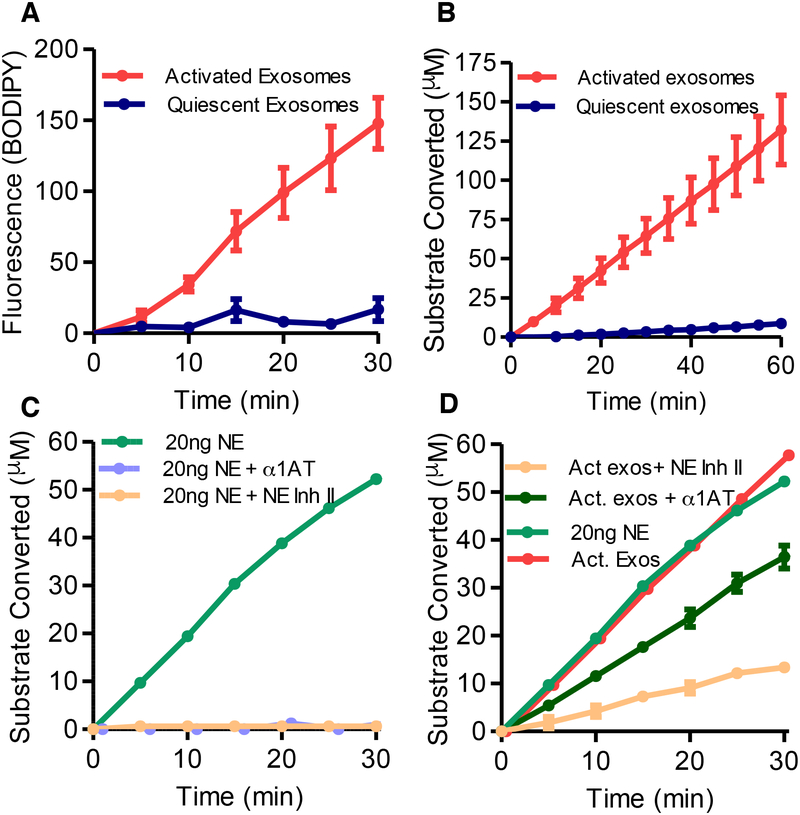
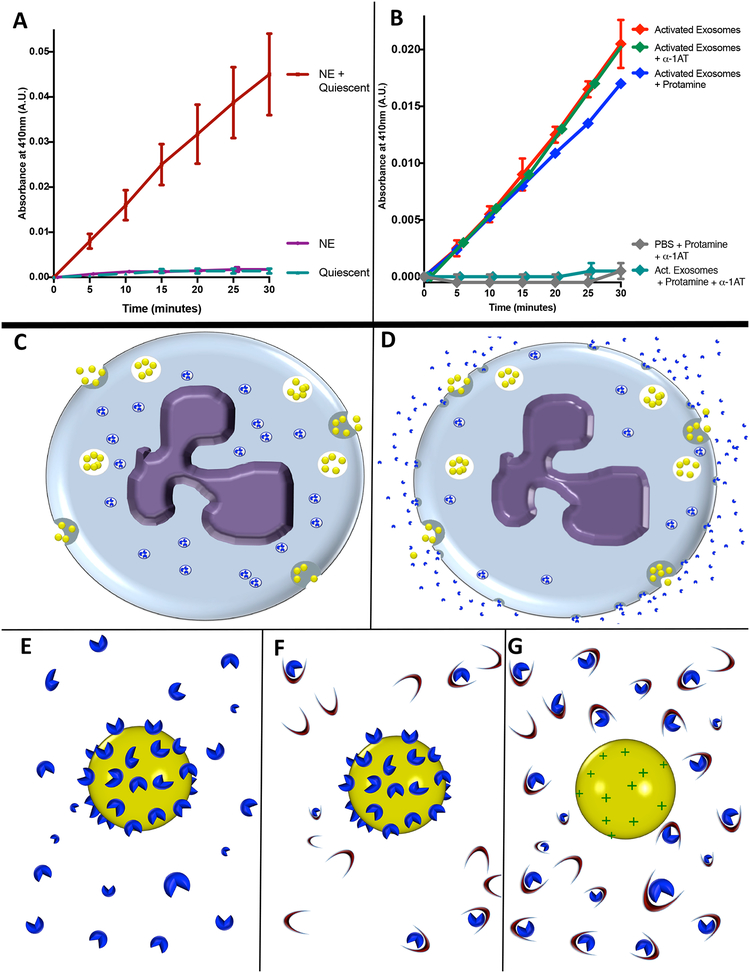
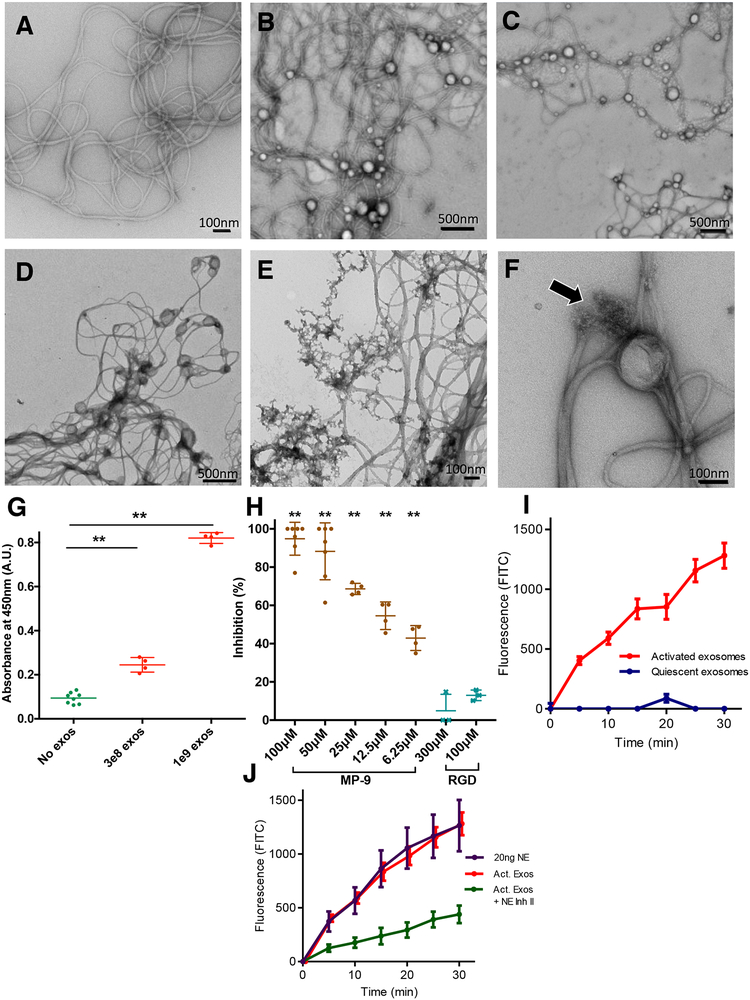
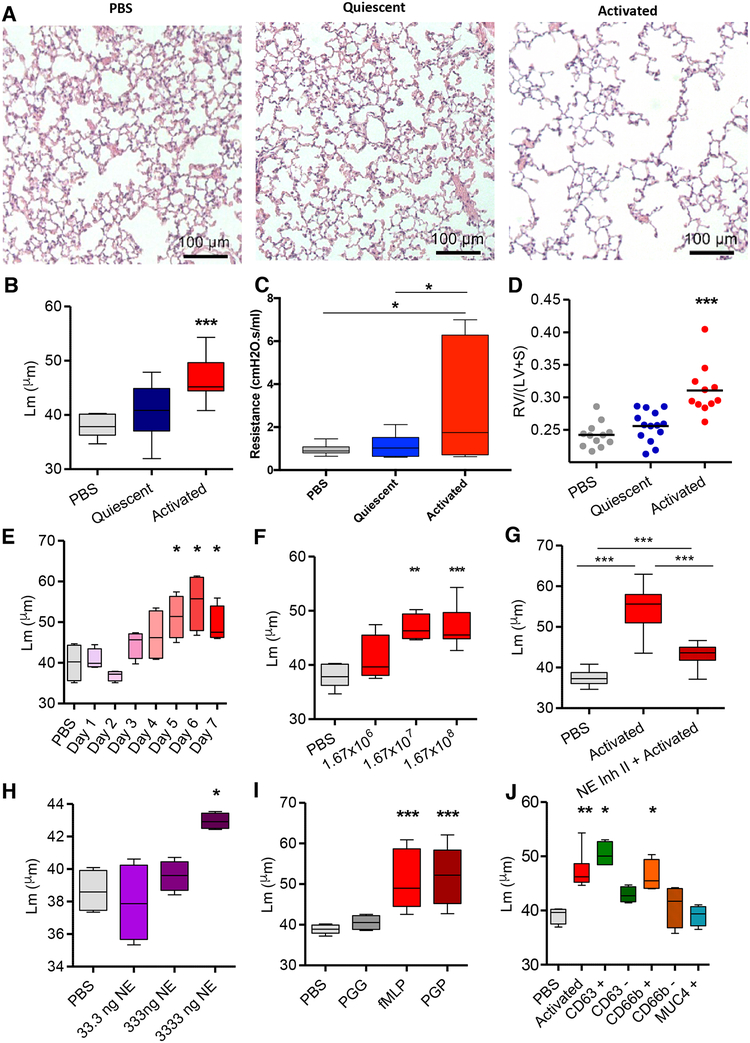
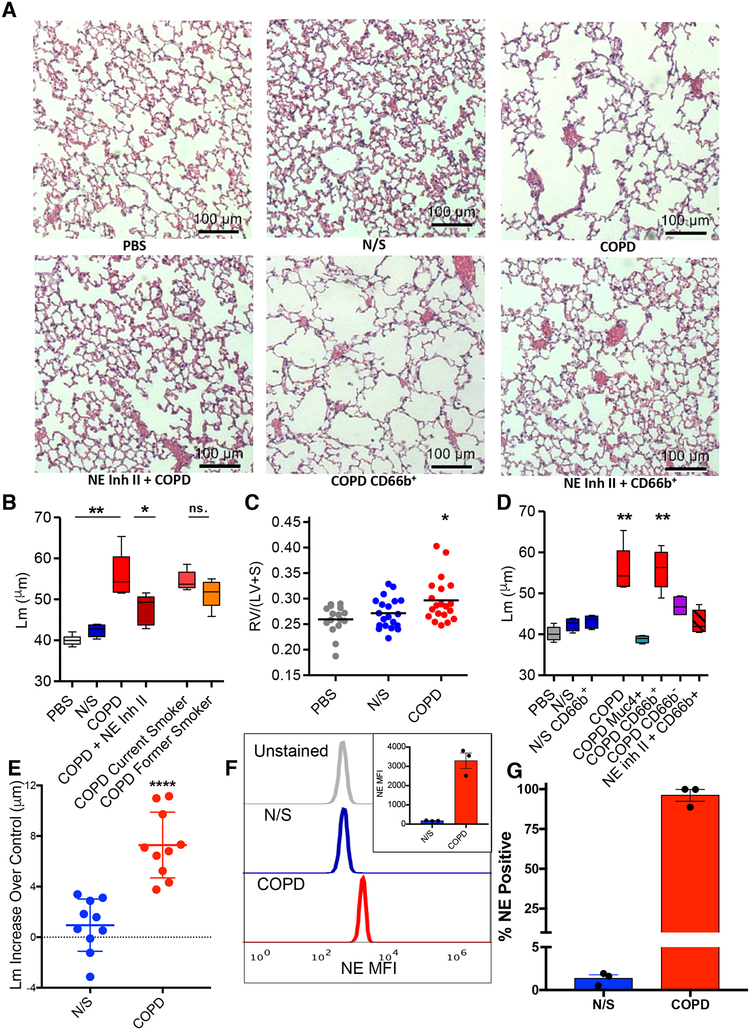
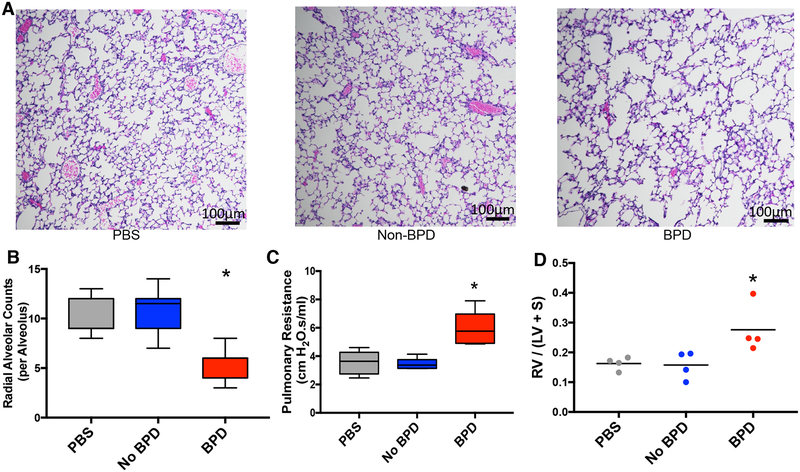
Here, we describe a novel pathogenic entity, the activated PMN (polymorphonuclear leukocyte, i.e., neutrophil)-derived exosome. These CD63+/CD66b+ nanovesicles acquire surface-bound neutrophil elastase (NE) during PMN degranulation, NE being oriented in a configuration resistant to α1-antitrypsin (α1AT). These exosomes bind and degrade extracellular matrix (ECM) via the integrin Mac-1 and NE, respectively, causing the hallmarks of chronic obstructive pulmonary disease (COPD). Due to both ECM targeting and α1AT resistance, exosomal NE is far more potent than free NE. Importantly, such PMN-derived exosomes exist in clinical specimens from subjects with COPD but not healthy controls and are capable of transferring a COPD-like phenotype from humans to mice in an NE-driven manner. Similar findings were observed for another neutrophil-driven disease of ECM remodeling (bronchopulmonary dysplasia [BPD]). These findings reveal an unappreciated role for exosomes in the pathogenesis of disorders of ECM homeostasis such as COPD and BPD, providing a critical mechanism for proteolytic damage.
Keywords: BPD; COPD; ELA-2; elastase; exosomes; extracellular matrix; extracellular vesicles; inflammation; lung disease; microparticles; neutrophil.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interest: The authors declare no competing interests.
Figures
Comment in
-
Recent Insights into In Utero Gene Editing, Checkpoint Inhibitors, and Polymorphonuclear Extracellular Vesicles.Am J Respir Cell Mol Biol. 2020 Jun;62(6):805-807. doi: 10.1165/rcmb.2020-0024RO. Am J Respir Cell Mol Biol. 2020. PMID: 32017596 No abstract available.
References
-
- Betsuyaku T, Nishimura M, Takeyabu K, Tanino M, Miyamoto K, and Kawakami Y (2000). Decline in FEV(1) in community-based older volunteers with higher levels of neutrophil elastase in bronchoalveolar lavage fluid. Respiration 67, 261–267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL114439/HL/NHLBI NIH HHS/United States
- I01 BX001756/BX/BLRD VA/United States
- K08 HL141652/HL/NHLBI NIH HHS/United States
- R01 HL102371/HL/NHLBI NIH HHS/United States
- K08 HL123940/HL/NHLBI NIH HHS/United States
- T32 HL105346/HL/NHLBI NIH HHS/United States
- R01 HL126596/HL/NHLBI NIH HHS/United States
- R35 HL135710/HL/NHLBI NIH HHS/United States
- P30 DK079337/DK/NIDDK NIH HHS/United States
- R01 HL126603/HL/NHLBI NIH HHS/United States
- R01 HL077783/HL/NHLBI NIH HHS/United States
- R01 HL110950/HL/NHLBI NIH HHS/United States
- P30 DK072482/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
