

Phage Lysis: Multiple Genes for Multiple Barriers
- PMID: 30635077
- PMCID: PMC6733033
- DOI: 10.1016/bs.aivir.2018.09.003
Phage Lysis: Multiple Genes for Multiple Barriers
Abstract
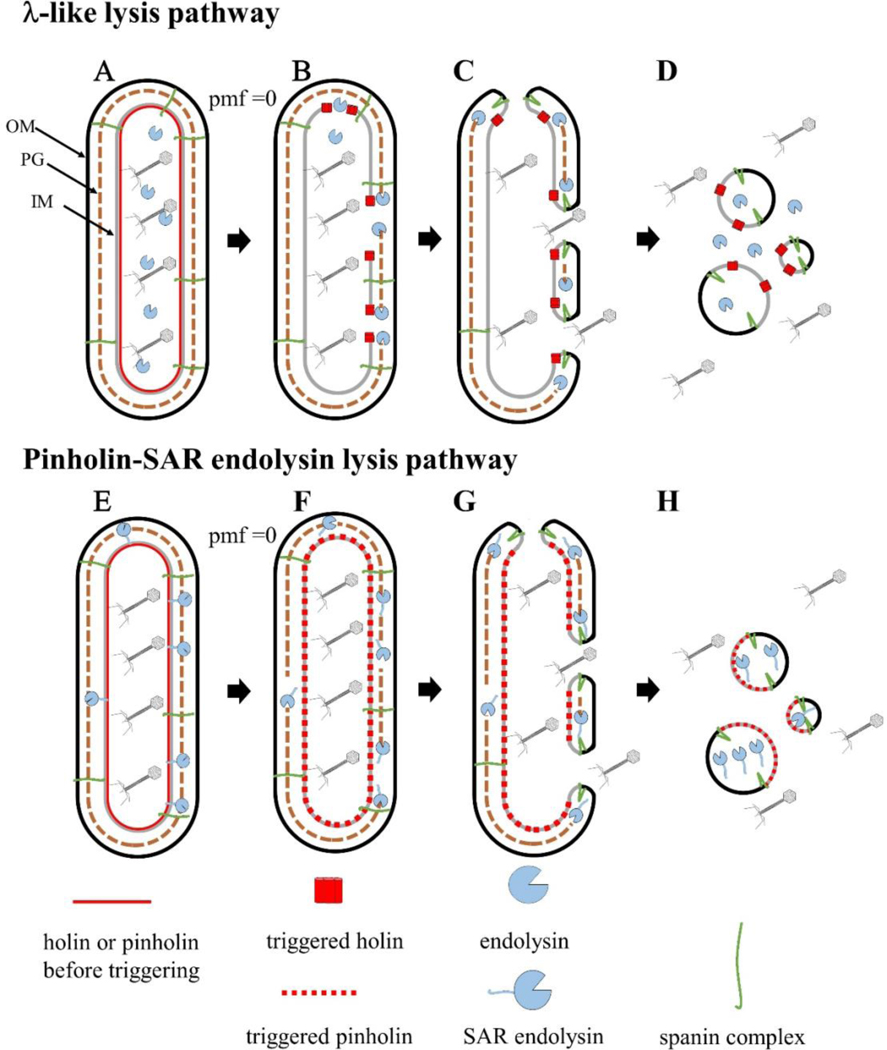
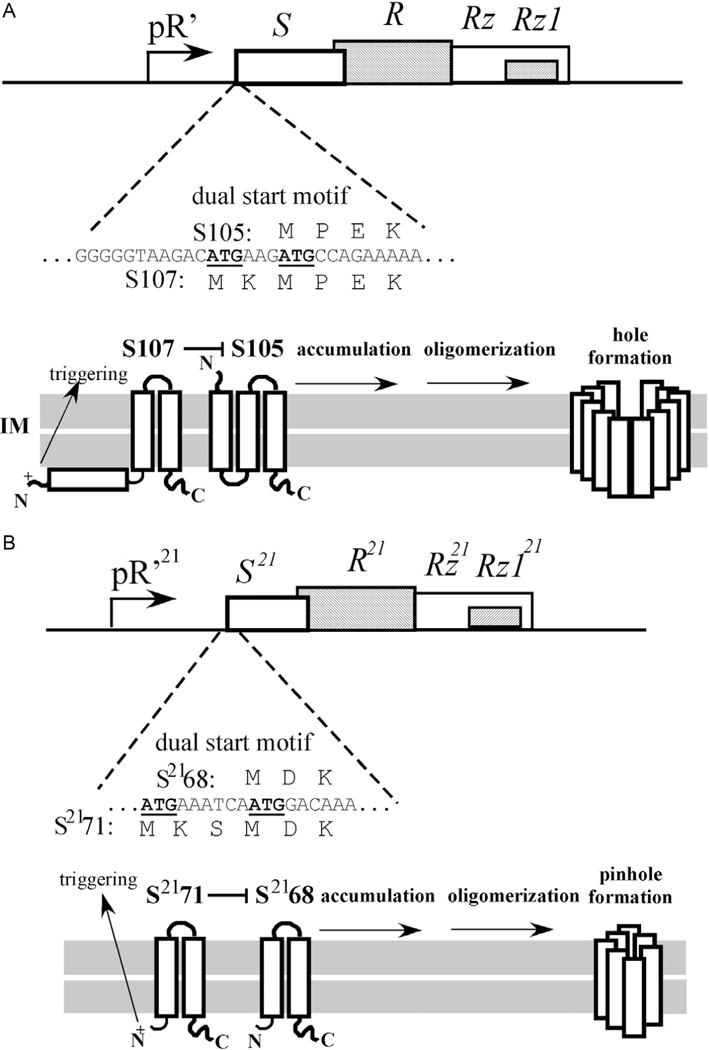
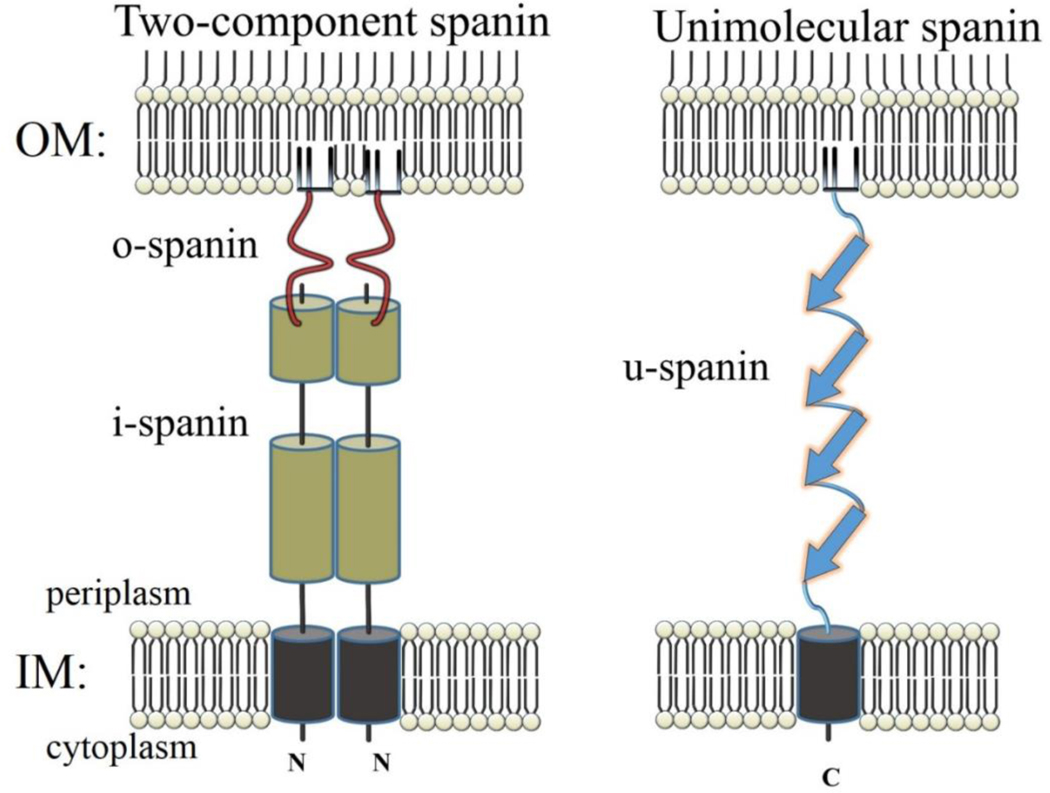
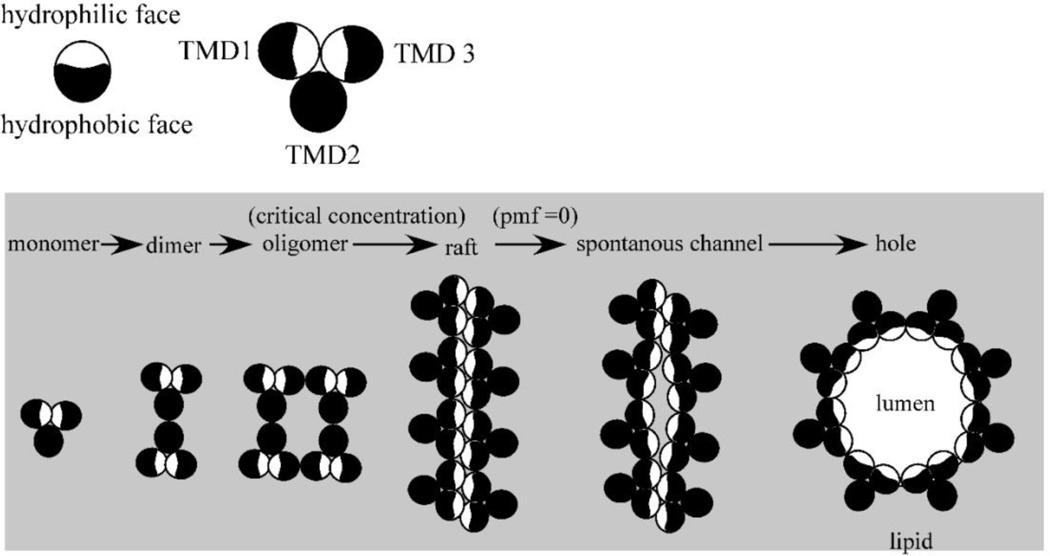
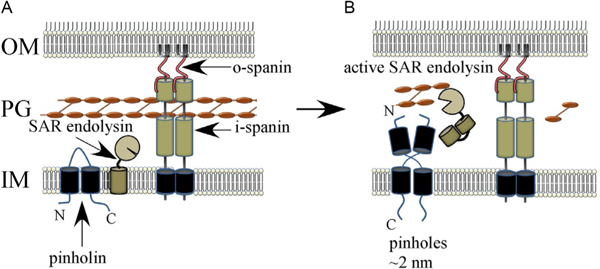
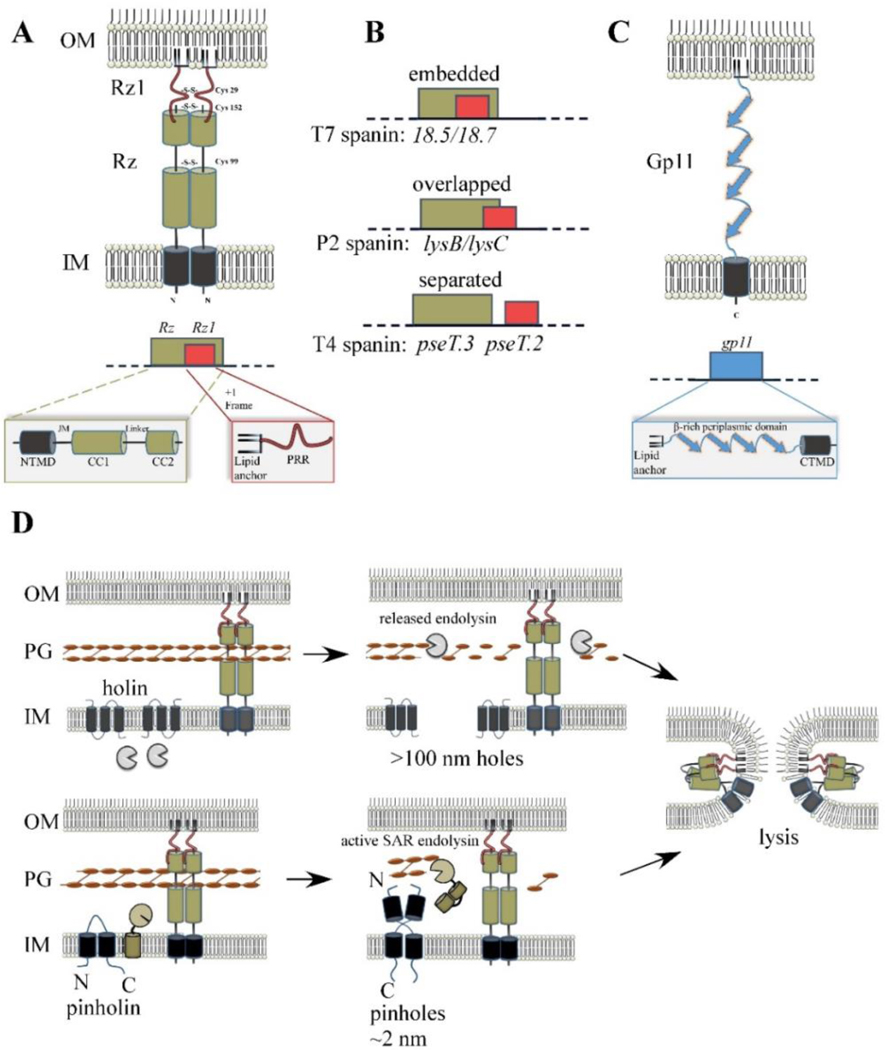
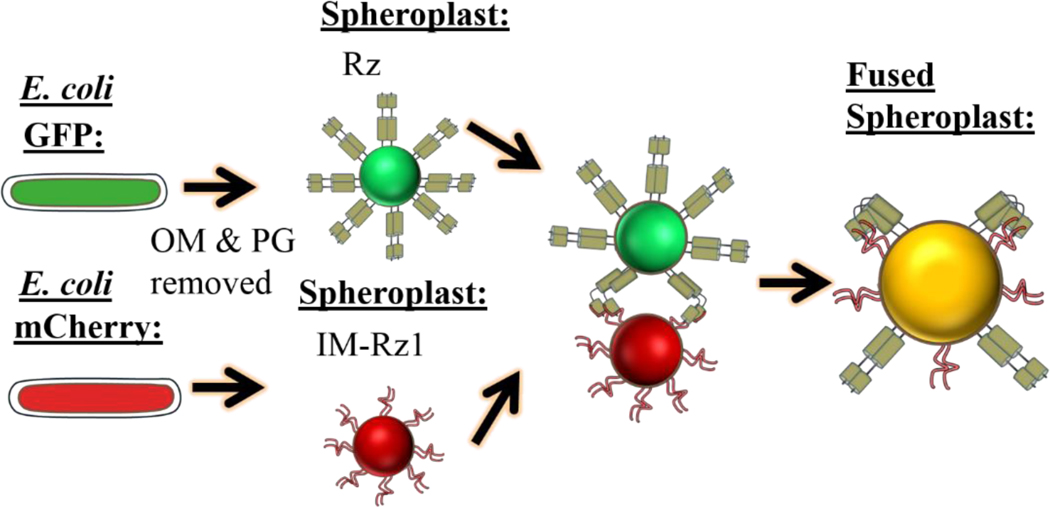
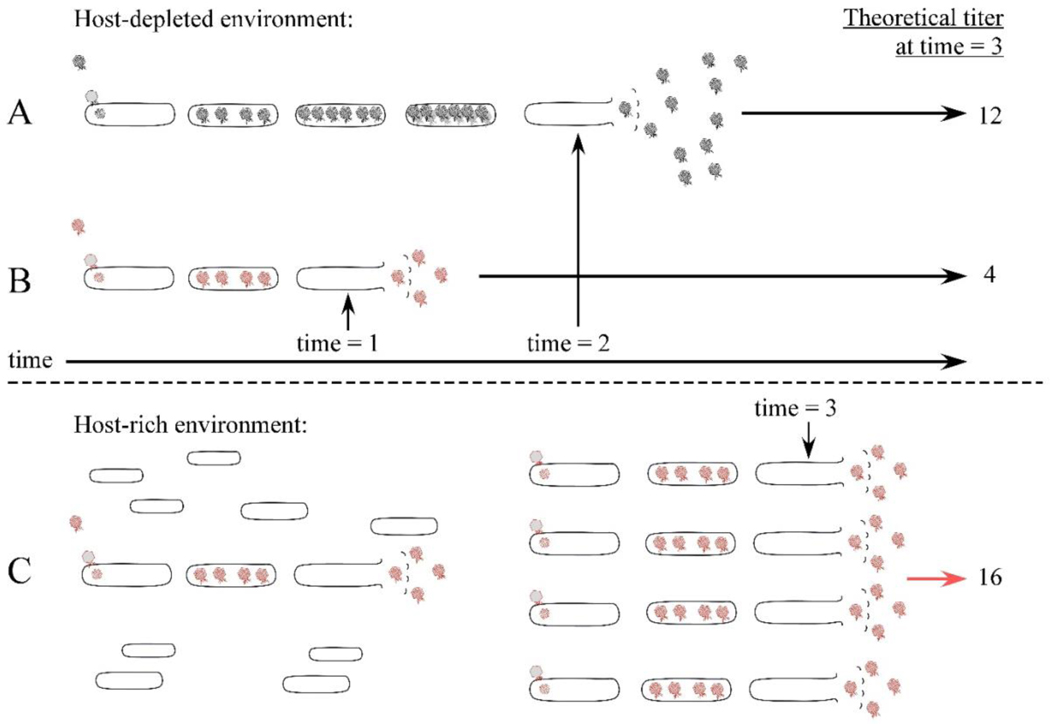
The first steps in phage lysis involve a temporally controlled permeabilization of the cytoplasmic membrane followed by enzymatic degradation of the peptidoglycan. For Caudovirales of Gram-negative hosts, there are two different systems: the holin-endolysin and pinholin-SAR endolysin pathways. In the former, lysis is initiated when the holin forms micron-scale holes in the inner membrane, releasing active endolysin into the periplasm to degrade the peptidoglycan. In the latter, lysis begins when the pinholin causes depolarization of the membrane, which activates the secreted SAR endolysin. Historically, the disruption of the first two barriers of the cell envelope was thought to be necessary and sufficient for lysis of Gram-negative hosts. However, recently a third functional class of lysis proteins, the spanins, has been shown to be required for outer membrane disruption. Spanins are so named because they form a protein bridge that connects both membranes. Most phages produce a two-component spanin complex, composed of an outer membrane lipoprotein (o-spanin) and an inner membrane protein (i-spanin) with a predominantly coiled-coil periplasmic domain. Some phages have a different type of spanin which spans the periplasm as a single molecule, by virtue of an N-terminal lipoprotein signal and a C-terminal transmembrane domain. Evidence is reviewed supporting a model in which the spanins function by fusing the inner membrane and outer membrane. Moreover, it is proposed that spanin function is inhibited by the meshwork of the peptidoglycan, thus coupling the spanin step to the first two steps mediated by the holin and endolysin.
Keywords: antiholin; endolysin; evolution; holin; lysis; multigene lysis; pinholin; single gene lysis; spanin.
© 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- Abedon ST, 1989. Selection for bacteriophage latent period length by bacterial density: a theoretical examination. Microb. Ecol. 18, 79. - PubMed
-
- Barenboim M, Chang CY, dib Hajj F, Young R, 1999. Characterization of the dual start motif of a class II holin gene. Mol. Microbiol. 32, 715. - PubMed
-
- Bass RB, Strop P, Barclay M, Rees DC, 2002. Crystal structure of Escherichia coli MscS, a voltage-modulated and mechanosensitive channel. Science 298, 1582. - PubMed
-
- Beremand MN, Blumenthal T, 1979. Overlapping genes in RNA phage: a new protein implicated in lysis. Cell 18, 257. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
