

Efficiency measures the conversion of agonist binding energy into receptor conformational change
- PMID: 30635369
- PMCID: PMC6445574
- DOI: 10.1085/jgp.201812215
Efficiency measures the conversion of agonist binding energy into receptor conformational change
Erratum in
-
Correction: Efficiency measures the conversion of agonist binding energy into receptor conformational change.J Gen Physiol. 2020 Jan 6;152(1):e20181221511252019c. doi: 10.1085/jgp.20181221511252019c. J Gen Physiol. 2020. PMID: 31816639 Free PMC article. No abstract available.
Abstract
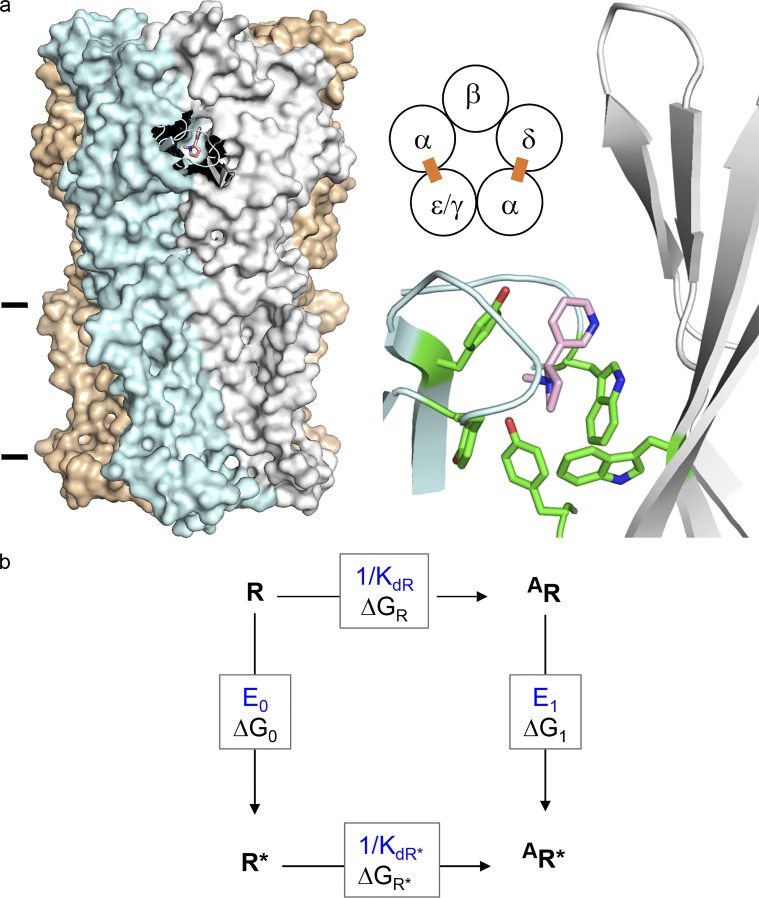
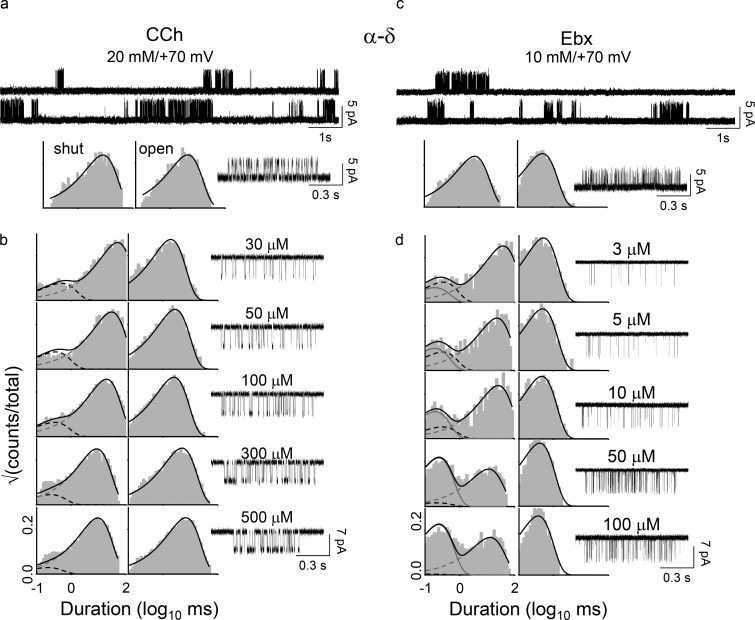
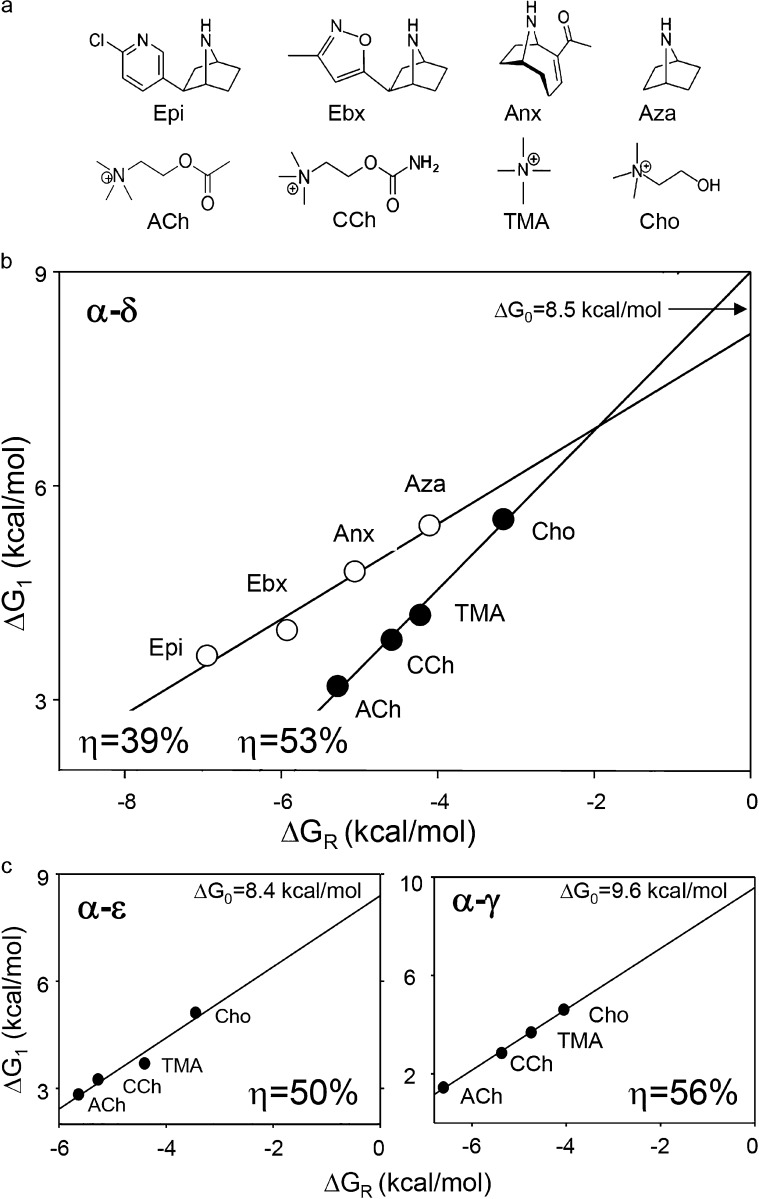
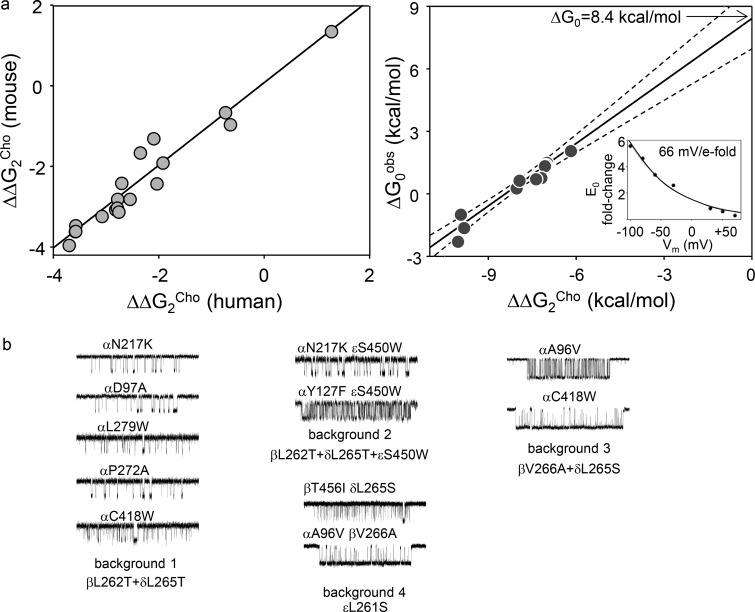
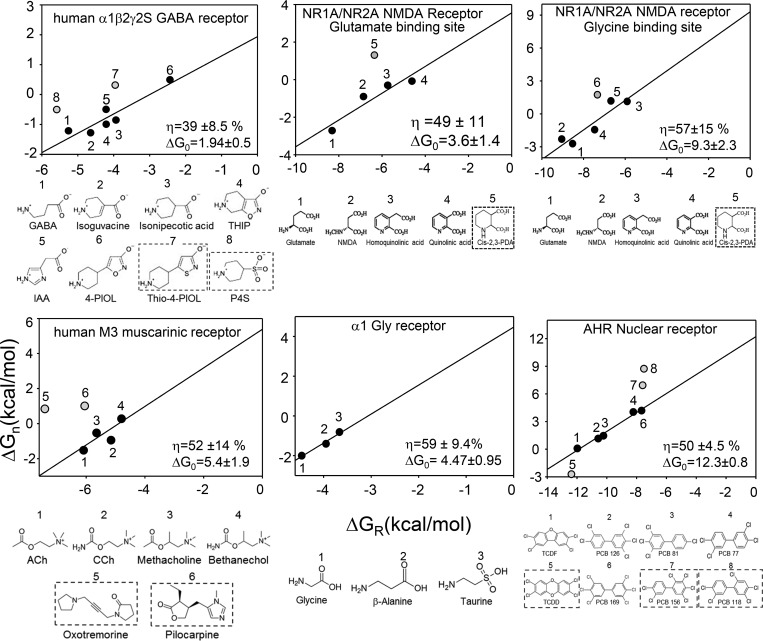
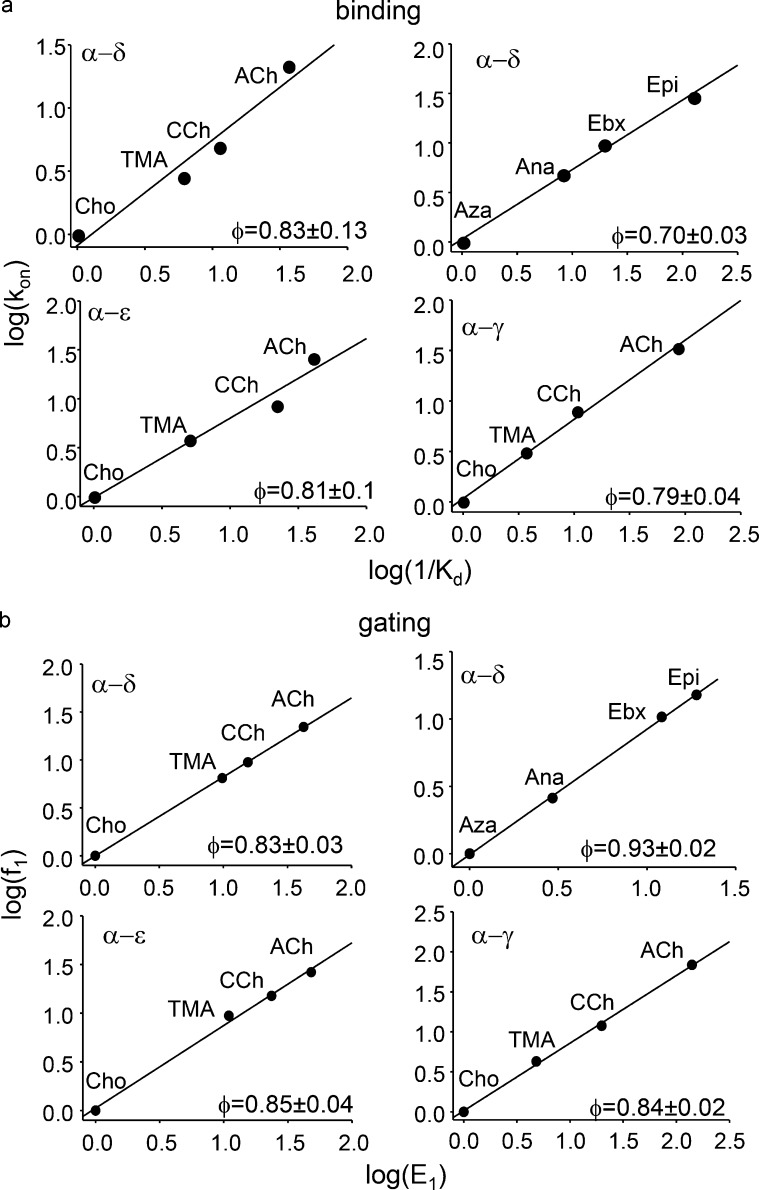
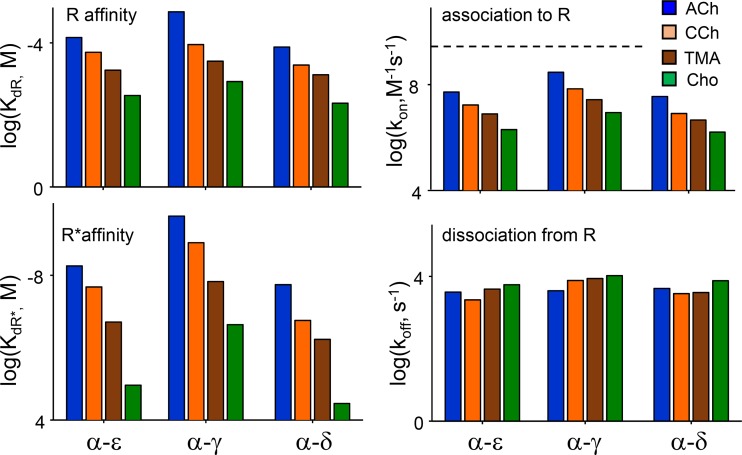
Receptors alternate between resting↔active conformations that bind agonists with low↔high affinity. Here, we define a new agonist attribute, energy efficiency (η), as the fraction of ligand-binding energy converted into the mechanical work of the activation conformational change. η depends only on the resting/active agonist-binding energy ratio. In a plot of activation energy versus binding energy (an "efficiency" plot), the slope gives η and the y intercept gives the receptor's intrinsic activation energy (without agonists; ΔG0). We used single-channel electrophysiology to estimate η for eight different agonists and ΔG0 in human endplate acetylcholine receptors (AChRs). From published equilibrium constants, we also estimated η for agonists of KCa1.1 (BK channels) and muscarinic, γ-aminobutyric acid, glutamate, glycine, and aryl-hydrocarbon receptors, and ΔG0 for all of these except KCa1.1. Regarding AChRs, η is 48-56% for agonists related structurally to acetylcholine but is only ∼39% for agonists related to epibatidine; ΔG0 is 8.4 kcal/mol in adult and 9.6 kcal/mol in fetal receptors. Efficiency plots for all of the above receptors are approximately linear, with η values between 12% and 57% and ΔG0 values between 2 and 12 kcal/mol. Efficiency appears to be a general attribute of agonist action at receptor binding sites that is useful for understanding binding mechanisms, categorizing agonists, and estimating concentration-response relationships.
© 2019 Nayak et al.
Figures
References
-
- Atkinson A. 1994. Fast very robust methods for the detection of multiple outliers. J. Am. Stat. Assoc. 89:1329–1339. 10.1080/01621459.1994.10476872 - DOI
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
