

F-box protein-32 down-regulates small-conductance calcium-activated potassium channel 2 in diabetic mouse atria
- PMID: 30635400
- PMCID: PMC6422073
- DOI: 10.1074/jbc.RA118.003837
F-box protein-32 down-regulates small-conductance calcium-activated potassium channel 2 in diabetic mouse atria
Abstract
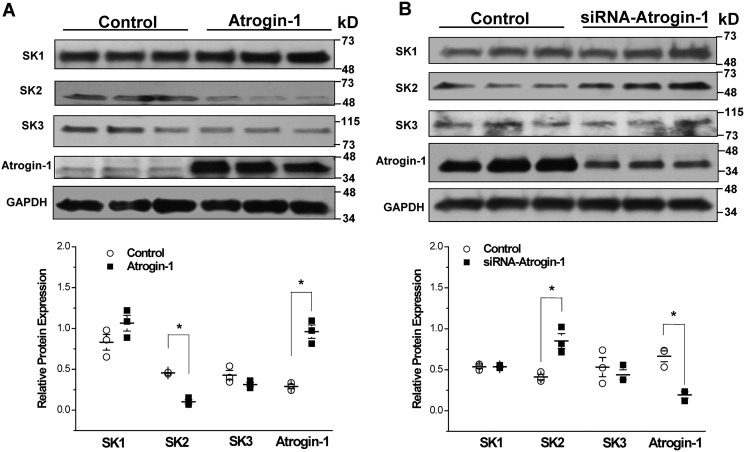
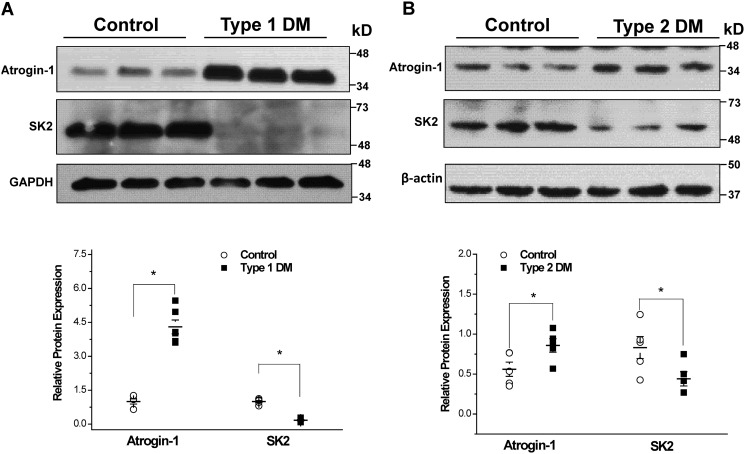
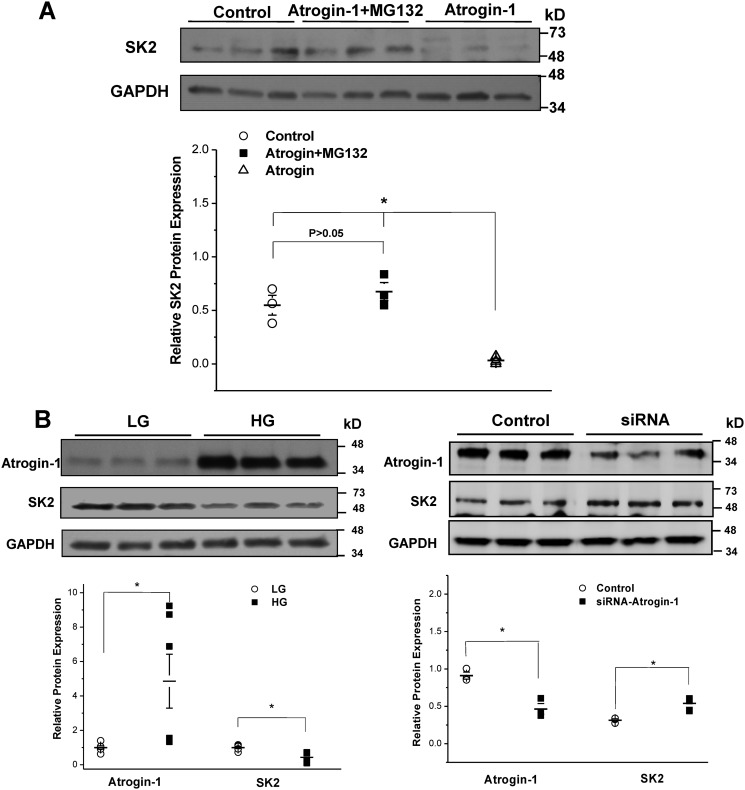
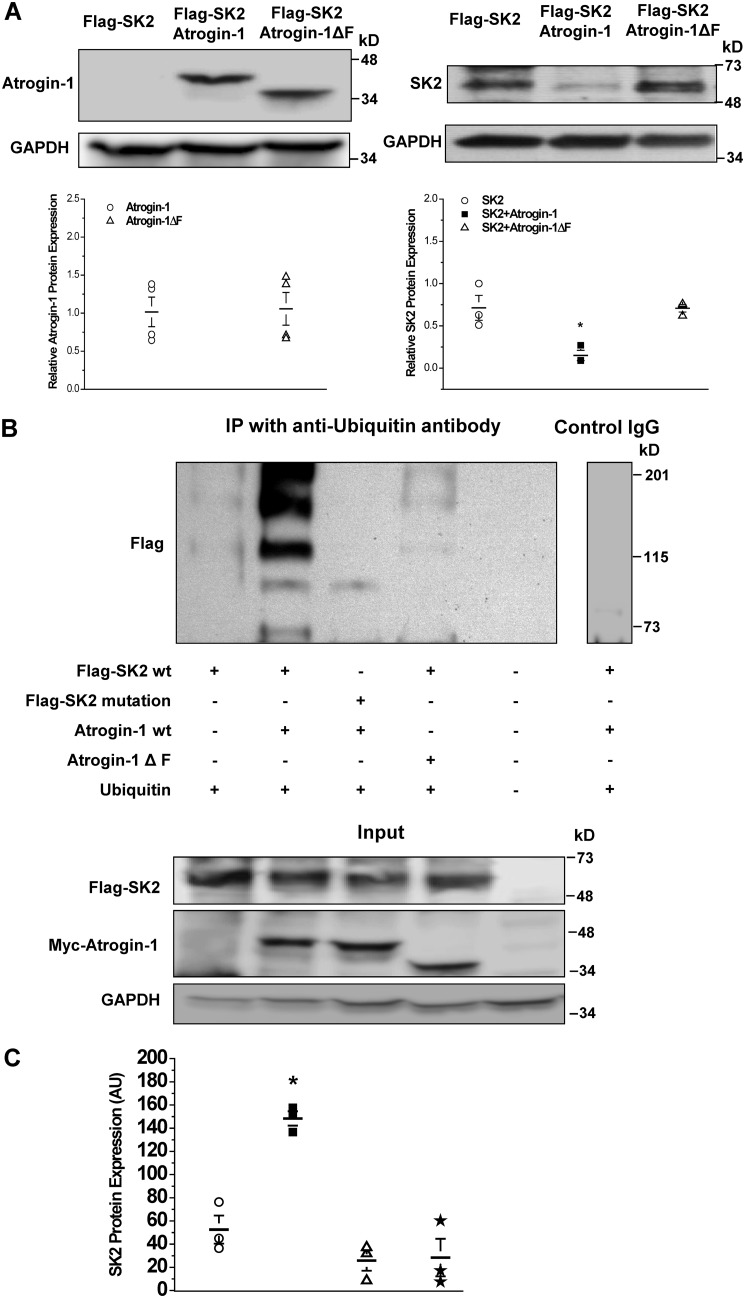
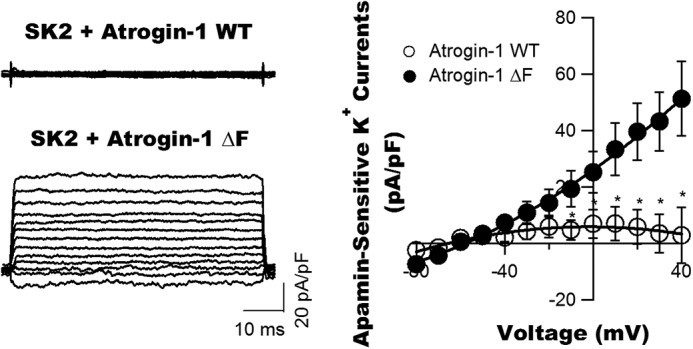
Diabetes mellitus (DM) is an independent risk factor for atrial fibrillation, but the underlying ionic mechanism for this association remains unclear. We recently reported that expression of the small-conductance calcium-activated potassium channel 2 (SK2, encoded by KCCN2) in atria from diabetic mice is significantly down-regulated, resulting in reduced SK currents in atrial myocytes from these mice. We also reported that the level of SK2 mRNA expression is not reduced in DM atria but that the ubiquitin-proteasome system (UPS), a major mechanism of intracellular protein degradation, is activated in vascular smooth muscle cells in DM. This suggests a possible role of the UPS in reduced SK currents. To test this possibility, we examined the role of the UPS in atrial SK2 down-regulation in DM. We found that a muscle-specific E3 ligase, F-box protein 32 (FBXO-32, also called atrogin-1), was significantly up-regulated in diabetic mouse atria. Enhanced FBXO-32 expression in atrial cells significantly reduced SK2 protein expression, and siRNA-mediated FBXO-32 knockdown increased SK2 protein expression. Furthermore, co-transfection of SK2 with FBXO-32 complementary DNA in HEK293 cells significantly reduced SK2 expression, whereas co-transfection with atrogin-1ΔF complementary DNA (a nonfunctional FBXO-32 variant in which the F-box domain is deleted) did not have any effects on SK2. These results indicate that FBXO-32 contributes to SK2 down-regulation and that the F-box domain is essential for FBXO-32 function. In conclusion, DM-induced SK2 channel down-regulation appears to be due to an FBXO-32-dependent increase in UPS-mediated SK2 protein degradation.
Keywords: HL-1 cells; SK channels; atrial fibrillation; atrogin-1; cardiomyocyte; cardiovascular disease; diabetes; diabetes mellitus; potassium channel; protein degradation; ubiquitin ligase.
© 2019 Ling et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health
Figures
Similar articles
-
ROS in diabetic atria regulate SK2 degradation by Atrogin-1 through the NF-κB signaling pathway.J Biol Chem. 2024 Mar;300(3):105735. doi: 10.1016/j.jbc.2024.105735. Epub 2024 Feb 8. J Biol Chem. 2024. PMID: 38336298 Free PMC article.
-
Muscle-specific f-box only proteins facilitate bk channel β(1) subunit downregulation in vascular smooth muscle cells of diabetes mellitus.Circ Res. 2010 Dec 10;107(12):1454-9. doi: 10.1161/CIRCRESAHA.110.228361. Epub 2010 Oct 21. Circ Res. 2010. PMID: 20966391 Free PMC article.
-
Down-regulation of the small conductance calcium-activated potassium channels in diabetic mouse atria.J Biol Chem. 2015 Mar 13;290(11):7016-26. doi: 10.1074/jbc.M114.607952. Epub 2015 Jan 20. J Biol Chem. 2015. PMID: 25605734 Free PMC article.
-
The role of E3 ubiquitin-ligases MuRF-1 and MAFbx in loss of skeletal muscle mass.Free Radic Biol Med. 2016 Sep;98:218-230. doi: 10.1016/j.freeradbiomed.2015.12.031. Epub 2015 Dec 29. Free Radic Biol Med. 2016. PMID: 26738803 Review.
-
The ubiquitin-proteasome system and skeletal muscle wasting.Essays Biochem. 2005;41:173-86. doi: 10.1042/EB0410173. Essays Biochem. 2005. PMID: 16250905 Review.
Cited by
-
Metabolic remodelling in atrial fibrillation: manifestations, mechanisms and clinical implications.Nat Rev Cardiol. 2024 Oct;21(10):682-700. doi: 10.1038/s41569-024-01038-6. Epub 2024 May 30. Nat Rev Cardiol. 2024. PMID: 38816507 Review.
-
The regulation of the small-conductance calcium-activated potassium current and the mechanisms of sex dimorphism in J wave syndrome.Pflugers Arch. 2021 Mar;473(3):491-506. doi: 10.1007/s00424-020-02500-3. Epub 2021 Jan 7. Pflugers Arch. 2021. PMID: 33411079 Free PMC article. Review.
-
FBXO32 promotes gastric cancer progression by regulating NME1.Transl Cancer Res. 2025 Jun 30;14(6):3520-3531. doi: 10.21037/tcr-2024-2426. Epub 2025 Jun 27. Transl Cancer Res. 2025. PMID: 40687258 Free PMC article.
-
Biobank-scale methods and projections for sparse polygenic prediction from machine learning.Sci Rep. 2023 Jul 19;13(1):11662. doi: 10.1038/s41598-023-37580-5. Sci Rep. 2023. PMID: 37468507 Free PMC article.
-
Genetic analysis reveals the shared genetic architecture between breast cancer and atrial fibrillation.Front Genet. 2025 Mar 25;16:1450259. doi: 10.3389/fgene.2025.1450259. eCollection 2025. Front Genet. 2025. PMID: 40201568 Free PMC article.
References
-
- World Health Organization (2016) Global report on diabetes. World Health Organization, Geneva, http://www.who.int/diabetes/global-report/en/
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
