

Allosteric disulfides: Sophisticated molecular structures enabling flexible protein regulation
- PMID: 30635401
- PMCID: PMC6393624
- DOI: 10.1074/jbc.REV118.005604
Allosteric disulfides: Sophisticated molecular structures enabling flexible protein regulation
Abstract
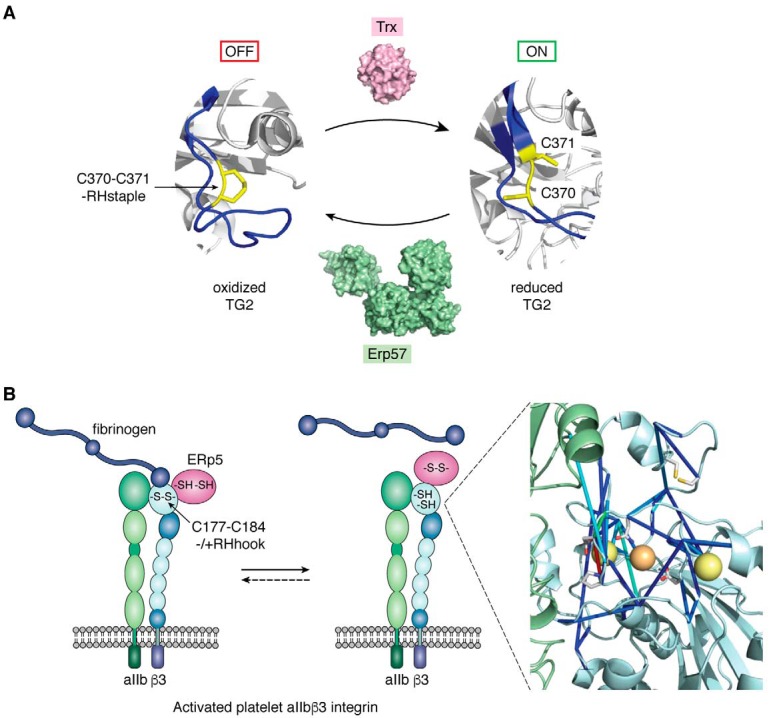
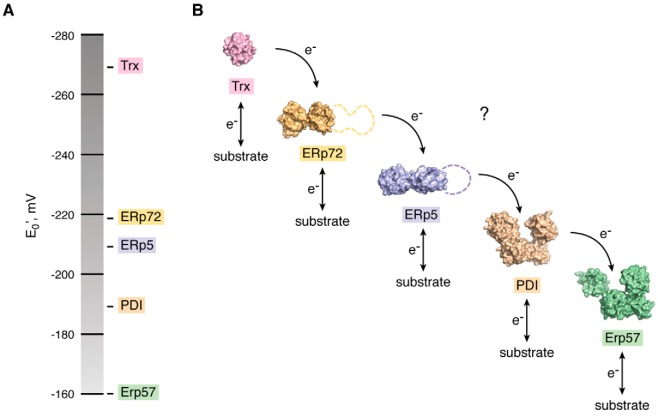
Protein disulfide bonds link pairs of cysteine residues in polypeptide chains. Many of these bonds serve a purely structural or energetic role, but a growing subset of cleavable disulfide bonds has been shown to control the function of the mature protein in which they reside. These allosteric disulfides and the factors that cleave these bonds are being identified across biological systems and life forms and have been shown to control hemostasis, the immune response, and viral infection in mammals. The discovery of these functional disulfides and a rationale for their facile nature has been aided by the emergence of a conformational signature for allosteric bonds. This post-translational modification mostly occurs extracellularly, making these chemical events prime drug targets. Indeed, a membrane-impermeable inhibitor of one of the cleaving factors is currently being trialed as an antithrombotic agent in cancer patients. Allosteric disulfides are firmly established as a sophisticated means by which a protein's shape and function can be altered; however, the full scope of this biological regulation will not be realized without new tools and techniques to study this regulation and innovative ways of targeting it.
Keywords: disulfide; protein chemistry; protein conformation; protein dynamic; protein disulfide isomerase; drug discovery; oxidation-reduction (redox); post-translational modification; protein chemical modification; thiol; allosteric; cysteine; cystine; disulfide bond; oxidoreductase.
© 2019 Chiu and Hogg.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures



References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
