

CNS-derived extracellular vesicles from superoxide dismutase 1 (SOD1)G93A ALS mice originate from astrocytes and neurons and carry misfolded SOD1
- PMID: 30635404
- PMCID: PMC6416428
- DOI: 10.1074/jbc.RA118.004825
CNS-derived extracellular vesicles from superoxide dismutase 1 (SOD1)G93A ALS mice originate from astrocytes and neurons and carry misfolded SOD1
Abstract
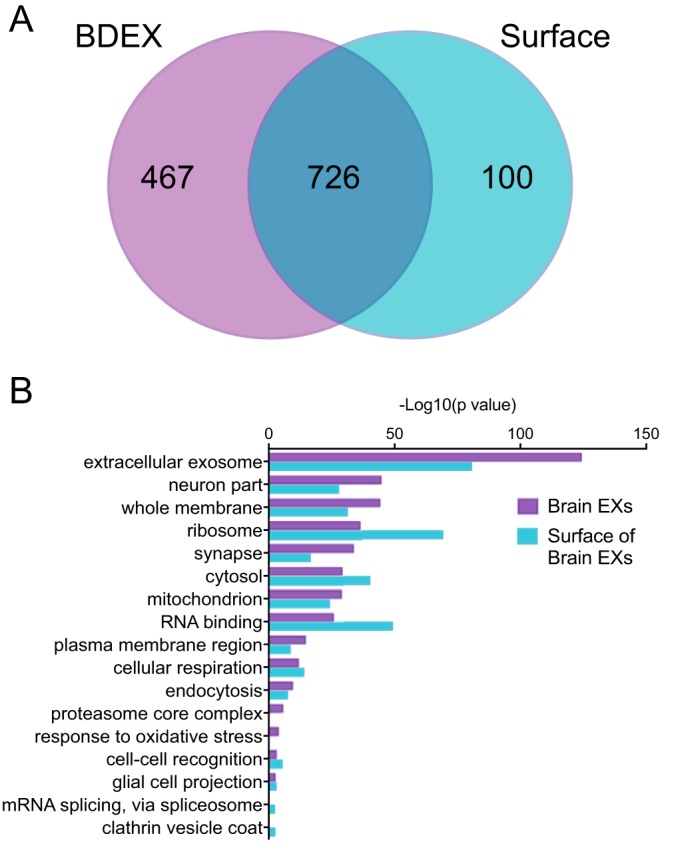
Extracellular vesicles (EVs) are secreted by myriad cells in culture and also by unicellular organisms, and their identification in mammalian fluids suggests that EV release also occurs at the organism level. However, although it is clearly important to better understand EVs' roles in organismal biology, EVs in solid tissues have received little attention. Here, we modified a protocol for EV isolation from primary neural cell culture to collect EVs from frozen whole murine and human neural tissues by serial centrifugation and purification on a sucrose gradient. Quantitative proteomics comparing brain-derived EVs from nontransgenic (NTg) and a transgenic amyotrophic lateral sclerosis (ALS) mouse model, superoxide dismutase 1 (SOD1)G93A, revealed that these EVs contain canonical exosomal markers and are enriched in synaptic and RNA-binding proteins. The compiled brain EV proteome contained numerous proteins implicated in ALS, and EVs from SOD1G93A mice were significantly depleted in myelin-oligodendrocyte glycoprotein compared with those from NTg animals. We observed that brain- and spinal cord-derived EVs, from NTg and SOD1G93A mice, are positive for the astrocyte marker GLAST and the synaptic marker SNAP25, whereas CD11b, a microglial marker, was largely absent. EVs from brains and spinal cords of the SOD1G93A ALS mouse model, as well as from human SOD1 familial ALS patient spinal cord, contained abundant misfolded and nonnative disulfide-cross-linked aggregated SOD1. Our results indicate that CNS-derived EVs from an ALS animal model contain pathogenic disease-causing proteins and suggest that brain astrocytes and neurons, but not microglia, are the main EV source.
Keywords: amyotrophic lateral sclerosis (ALS) (Lou Gehrig disease); astrocyte; central nervous system (CNS); exosome (vesicle); extracellular vesicles; neurodegeneration; protein homeostasis; proteomics; secretion.
© 2019 Silverman et al.
Conflict of interest statement
N. R. C. is the Chief Scientific Officer of ProMIS Neurosciences, which has licensed the 3H1 misfolded SOD1–specific antibody technology
Figures






References
-
- Grad L. I., Guest W. C., Yanai A., Pokrishevsky E., O'Neill M. A., Gibbs E., Semenchenko V., Yousefi M., Wishart D. S., Plotkin S. S., and Cashman N. R. (2011) Intermolecular transmission of superoxide dismutase 1 misfolding in living cells. Proc. Natl. Acad. Sci. U.S.A. 108, 16398–16403 10.1073/pnas.1102645108 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
