

A Nervous System-Specific Model of Creatine Transporter Deficiency Recapitulates the Cognitive Endophenotype of the Disease: a Longitudinal Study
- PMID: 30635645
- PMCID: PMC6329805
- DOI: 10.1038/s41598-018-37303-1
A Nervous System-Specific Model of Creatine Transporter Deficiency Recapitulates the Cognitive Endophenotype of the Disease: a Longitudinal Study
Abstract
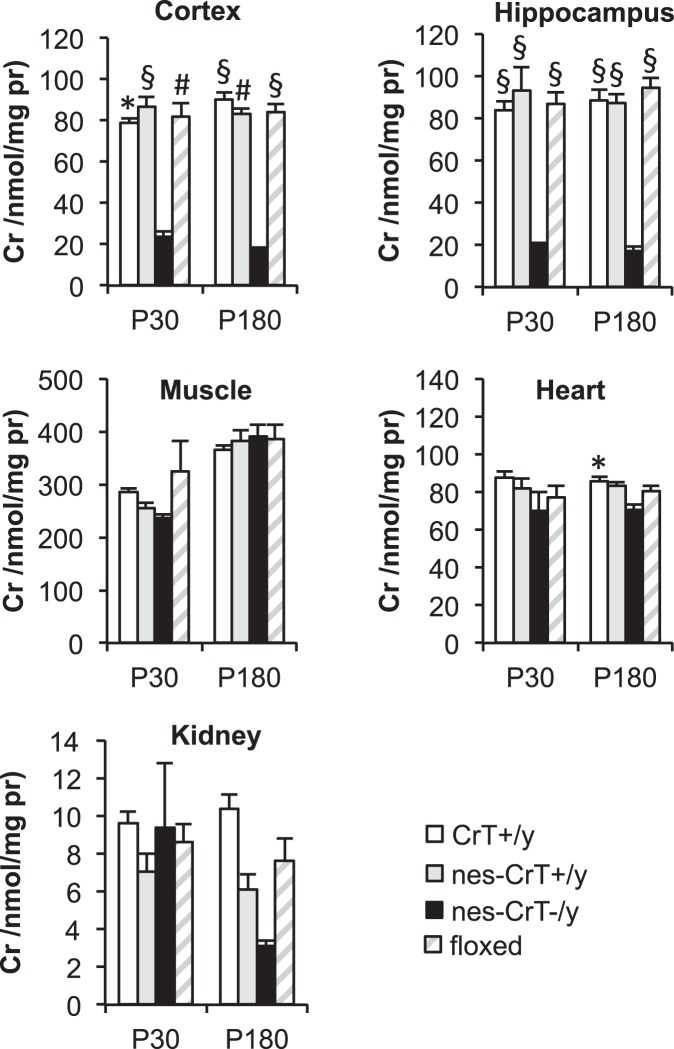
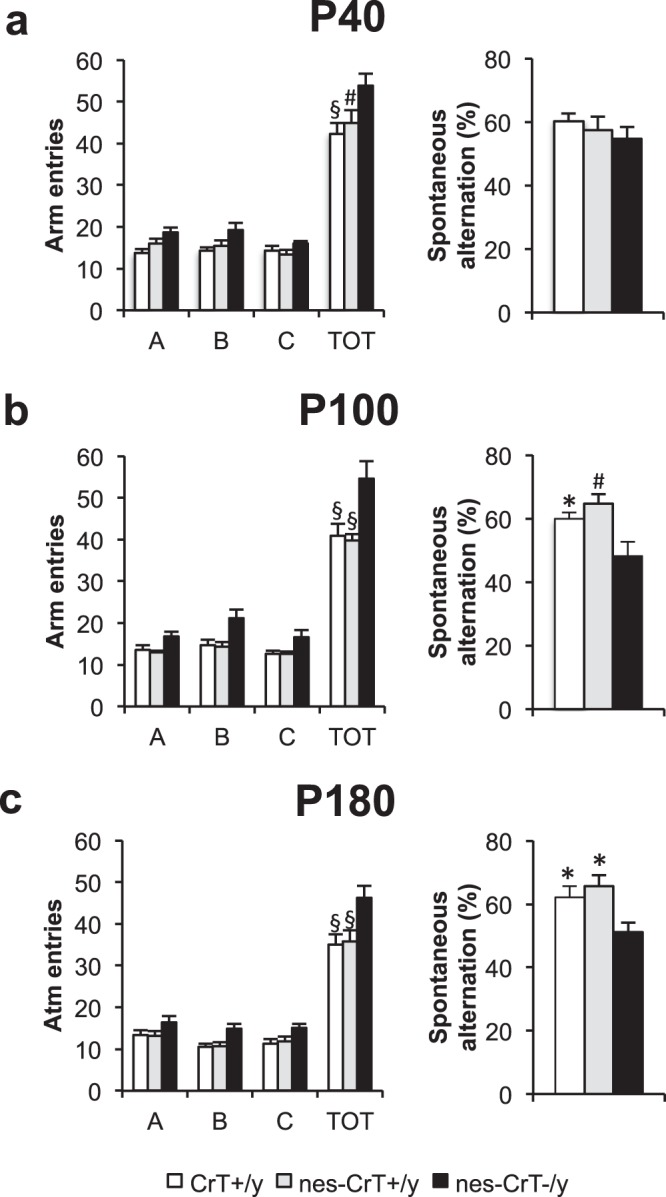
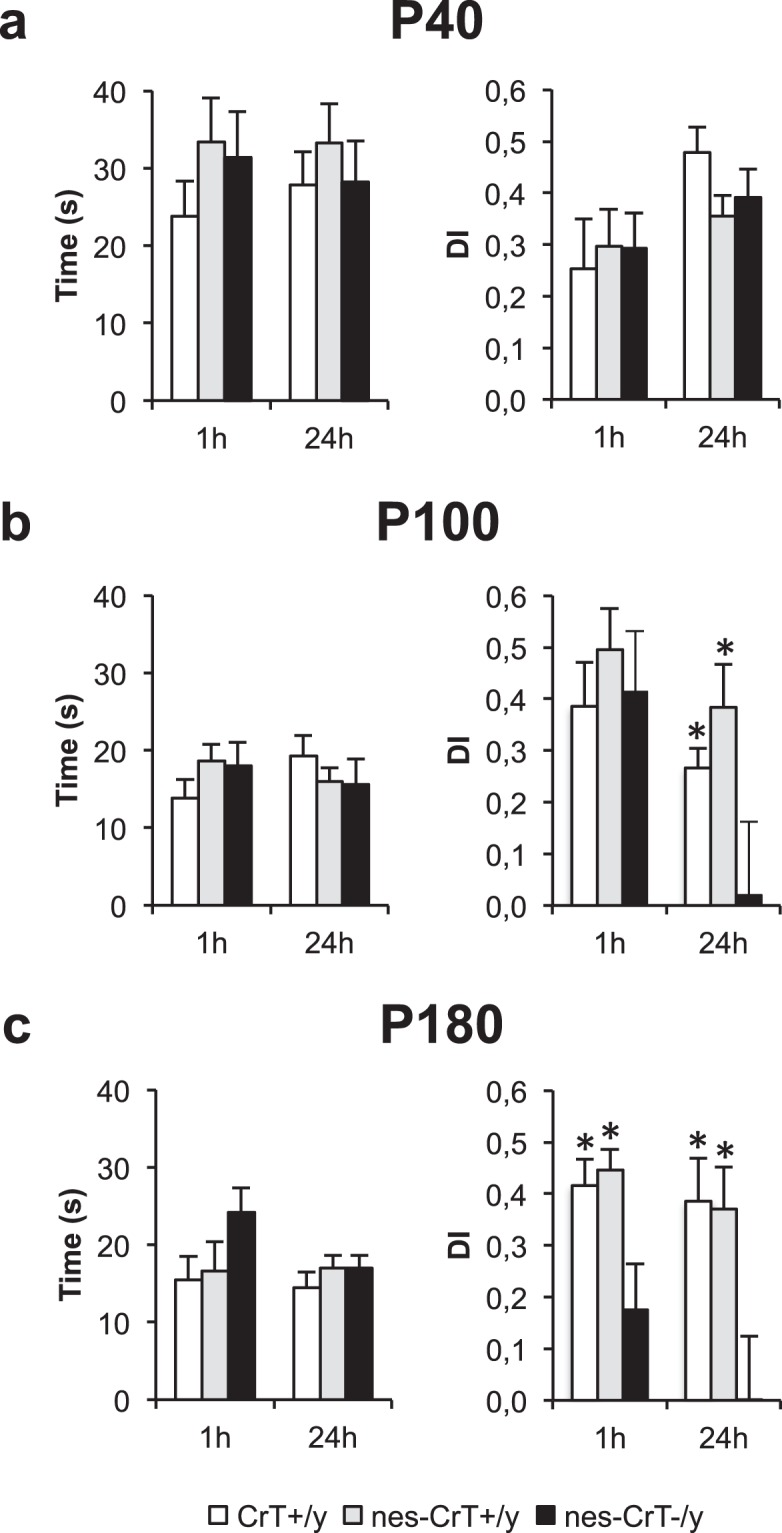
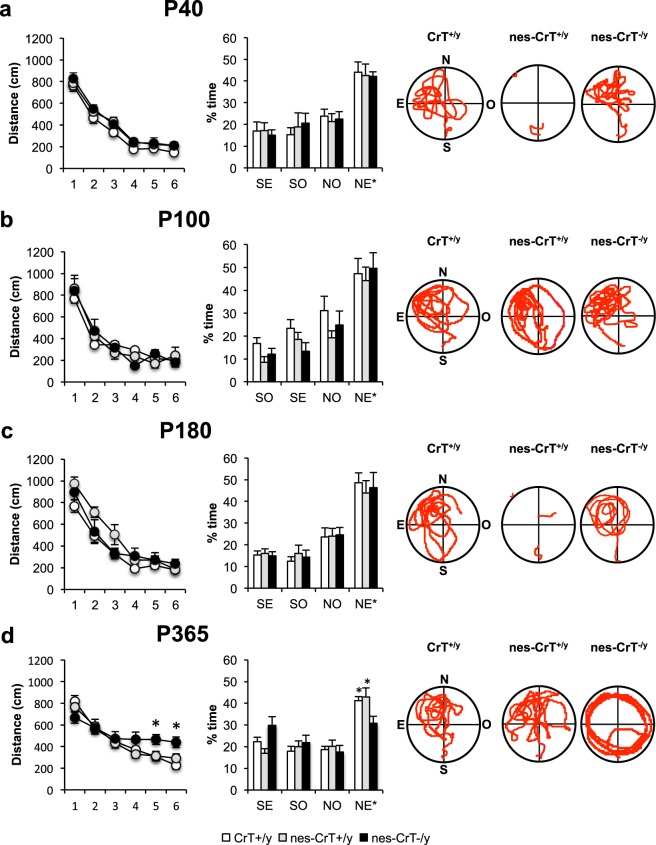
Mutations in creatine (Cr) transporter (CrT) gene lead to cerebral creatine deficiency syndrome-1 (CTD), an orphan neurodevelopmental disorder presenting with brain Cr deficiency, intellectual disability, seizures, movement and autistic-like behavioral disturbances, language and speech impairment. We have recently generated a murine model of CTD obtained by ubiquitous deletion of 5-7 exons in the CrT gene. These mice showed a marked Cr depletion, associated to early and progressive cognitive impairment, and autistic-like defects, thus resembling the key features of human CTD. Given the importance of extraneural dysfunctions in neurodevelopmental disorders, here we analyzed the specific role of neural Cr in the CTD phenotype. We induced the conditional deletion of Slc6a8 gene in neuronal and glial cells by crossing CrT floxed mice with the Nestin::Cre recombinase Tg (Nes-cre) 1Kln mouse. We report that nervous system-specific Cr depletion leads to a progressive cognitive regression starting in the adult age. No autistic-like features, including repetitive and stereotyped movements, routines and rituals, are present in this model. These results indicate that Cr depletion in the nervous system is a pivotal cause of the CTD pathological phenotype, in particular with regard to the cognitive domain, but extraneural actors also play a role.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
