

Adenosine heteroreceptor complexes in the basal ganglia are implicated in Parkinson's disease and its treatment
- PMID: 30637481
- PMCID: PMC6456481
- DOI: 10.1007/s00702-019-01969-2
Adenosine heteroreceptor complexes in the basal ganglia are implicated in Parkinson's disease and its treatment
Abstract
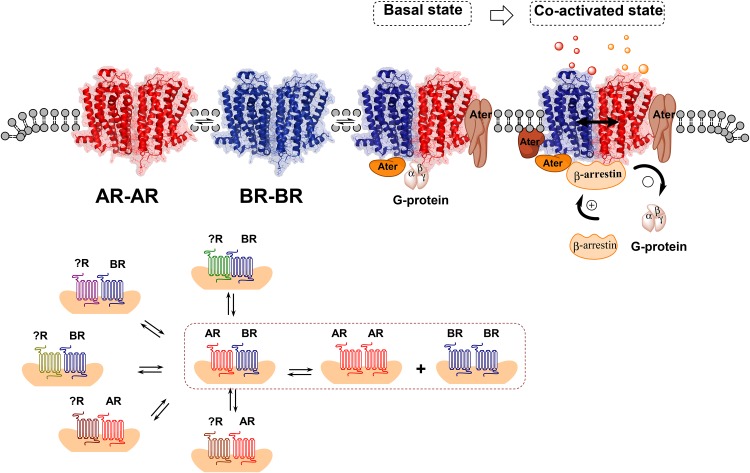
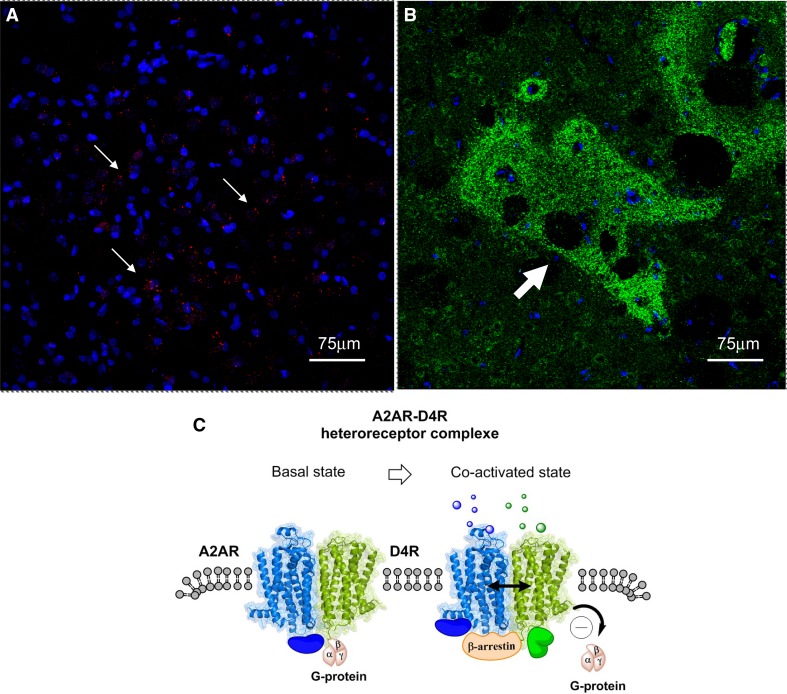
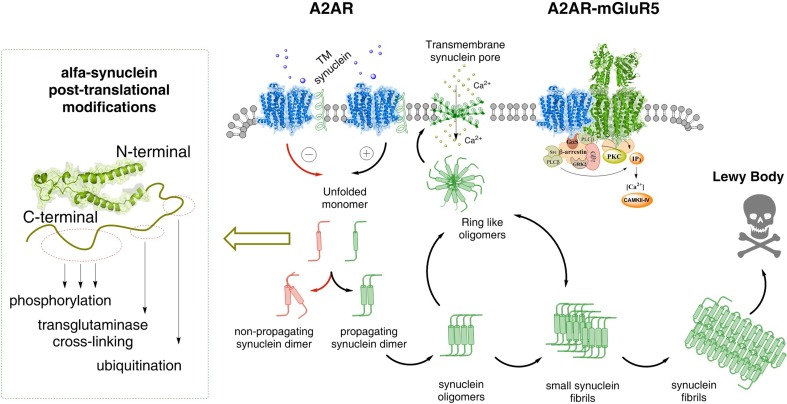
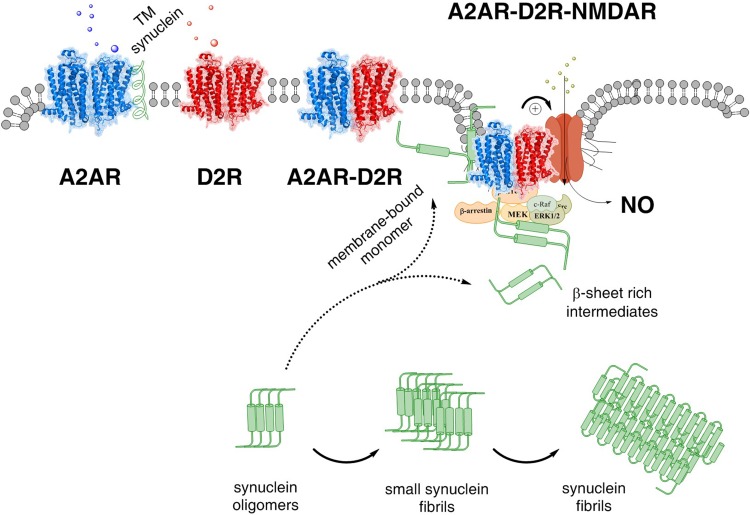
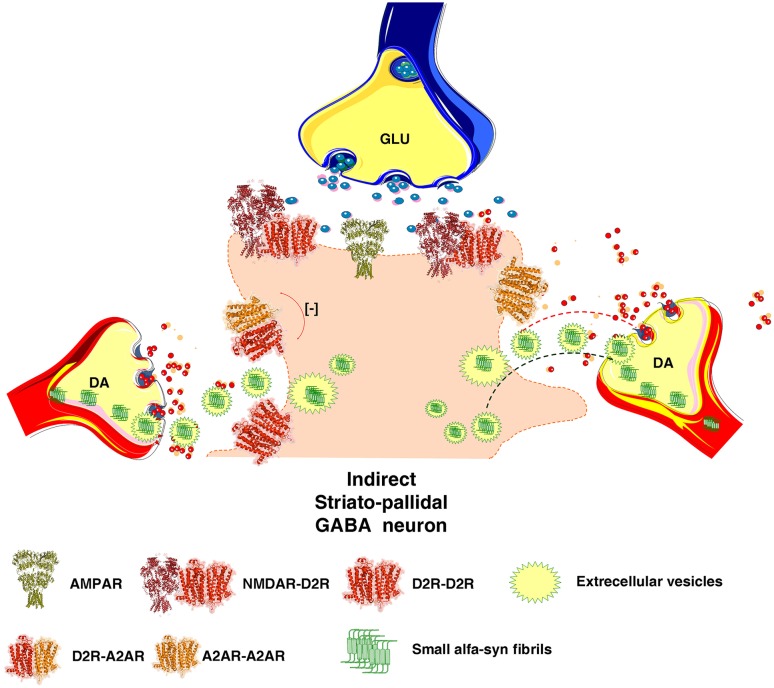
The adenosine homo, iso and heteroreceptor complexes in the basal ganglia play a highly significant role in modulating the indirect and direct pathways and the striosomal projections to the nigro-striatal DA system. The major adenosine receptor complexes in the striato-pallidal GABA neurons can be the A2AR-D2R and A2AR-D2R-mGluR5 receptor complexes, in which A2AR protomers and mGluR5 protomers can allosterically interact to inhibit D2R protomer signaling. Through a reorganization of these heteroreceptor complexes upon chronic dopaminergic treatment a pathological and prolonged inhibition of D2R receptor protomer signaling can develop with motor inhibition and wearing off of the therapeutic effects of levodopa and dopamine receptor agonists. The direct pathway is enriched in D1R in and around glutamate synapses enhancing the ability of these GABA neurons to be activated and increase motor initiation. The brake on these GABA neurons is in this case exerted by A1R forming A1R-D1R heteroreceptor complexes in which they allosterically inhibit D1R signaling and thereby reduce motor initiation. Upon chronic levodopa treatment a reorganization of the D1R heteroreceptor complexes develops with the formation of putative A1R-D1R-D3 in addition to D1R-D3R complexes in which D3R enhances D1R protomer signaling and may make the A1R protomer brake less effective. Alpha-synuclein monomers-dimers are postulated to form complexes with A2AR homo and heteroprotomers in the plasma membrane enhancing alpha-synuclein aggregation and toxicity. The alpha-synuclein fibrils formed in the A2AR enriched dendritic spines of the striato-pallidal GABA neurons may reach the surrounding DA terminals via extracellular-vesicle-mediated volume transmission involving internalization of the vesicles and their cargo (alpha-synuclein fibrils) into the vulnerable DA terminals, enhancing their degeneration followed by retrograde flow of these fibrils in the DA axons to the vulnerable nigral DA nerve cells.
Keywords: Adenosine receptor; Basal ganglia; G protein-coupled receptor; Heteroreceptor complexes; Neurodegeneration; Oligomerization; Parkinson’s diseases.
Figures
References
-
- Anden NE, Carlsson A, Dahlstroem A, Fuxe K, Hillarp NA, Larsson K. Demonstration and mapping out of nigro-neoestriatal dopamine neurons. Life Sci. 1964;3:523–530. - PubMed
-
- Antonelli T, Fuxe K, Agnati L, Mazzoni E, Tanganelli S, Tomasini MC, Ferraro L. Experimental studies and theoretical aspects on A2A/D2 receptor interactions in a model of Parkinson’s disease. relevance for L-dopa induced dyskinesias. J Neurol Sci. 2006;248:16–22. - PubMed
-
- Audet N, Galés C, Archer-Lahlou E, Vallières M, Schiller PW, Bouvier M, Pineyro G. Bioluminescence resonance energy transfer assays reveal ligand-specific conformational changes within preformed signaling complexes containing delta-opioid receptors and heterotrimeric G proteins. J Biol Chem. 2008;283:15078–15088. - PMC - PubMed
-
- Azdad K, Gall D, Woods AS, Ledent C, Ferre S, Schiffmann SN. Dopamine D2 and adenosine A2A receptors regulate NMDA-mediated excitation in accumbens neurons through A2A-D2 receptor heteromerization Neuropsychopharmacology: official publication of the American College. Neuropsychopharmacology. 2009;34:972–986. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
