

DNP-Assisted NMR Investigation of Proteins at Endogenous Levels in Cellular Milieu
- PMID: 30638534
- PMCID: PMC8982140
- DOI: 10.1016/bs.mie.2018.08.023
DNP-Assisted NMR Investigation of Proteins at Endogenous Levels in Cellular Milieu
Abstract
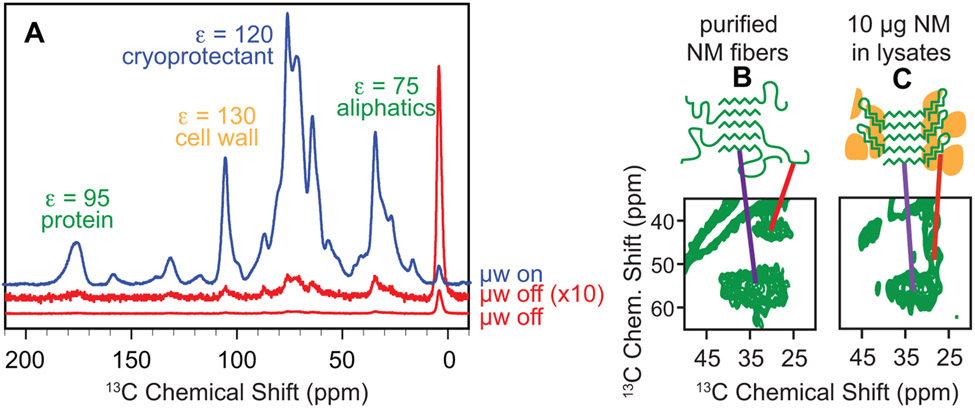
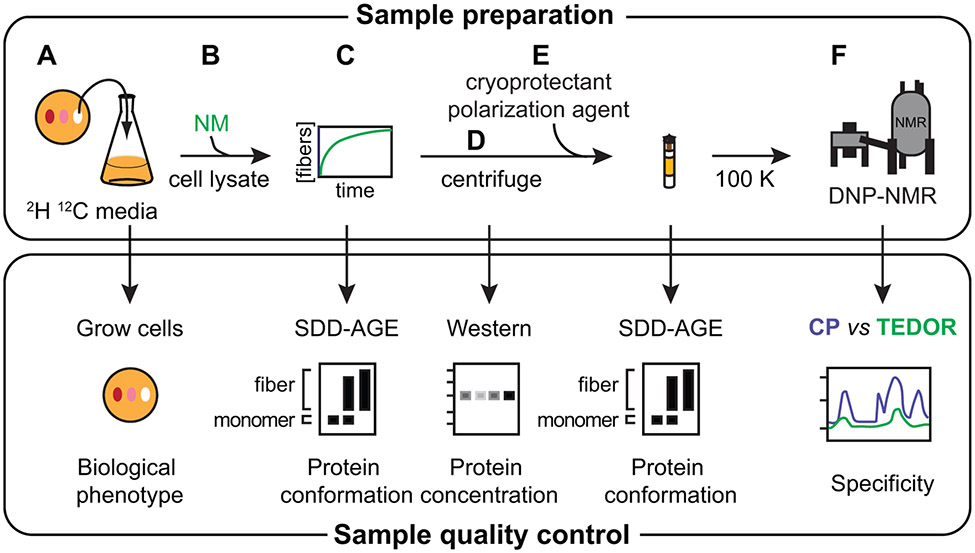
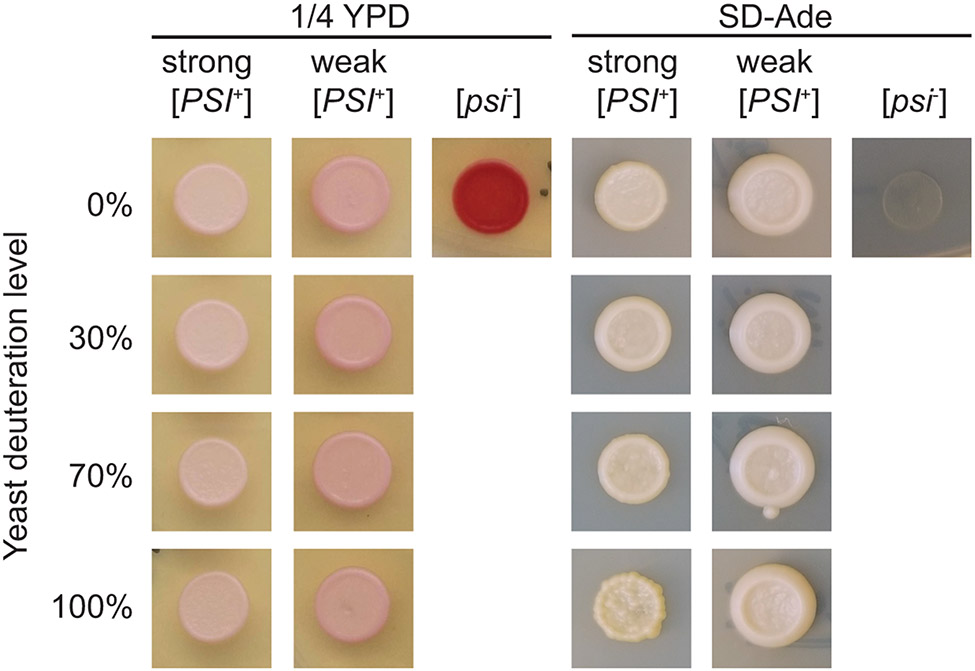
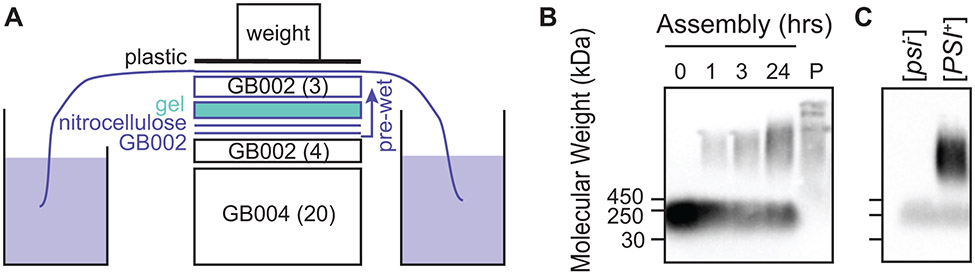
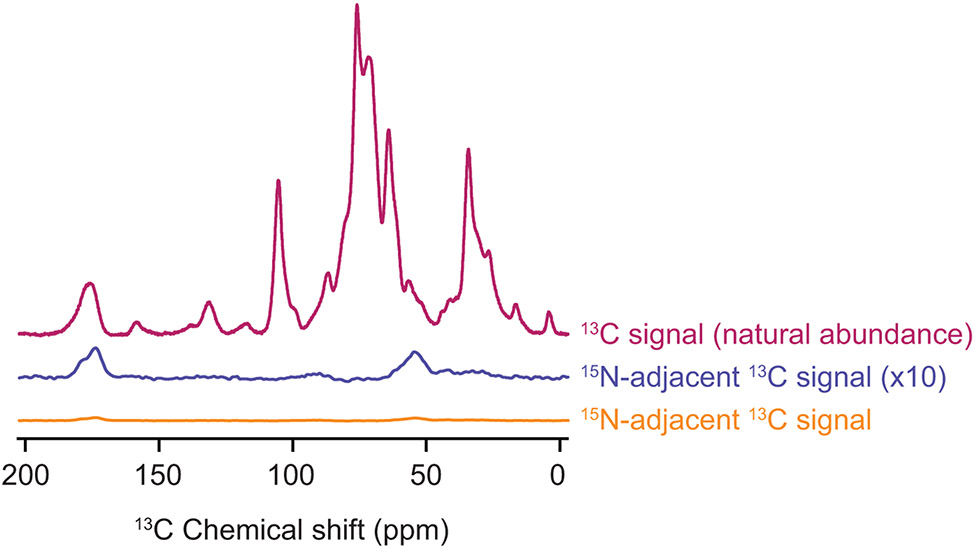
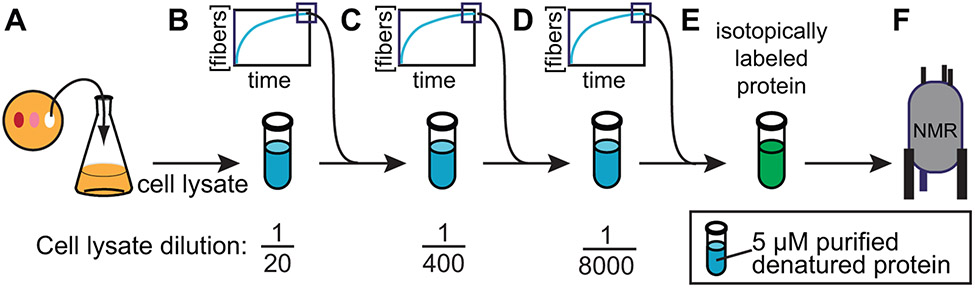
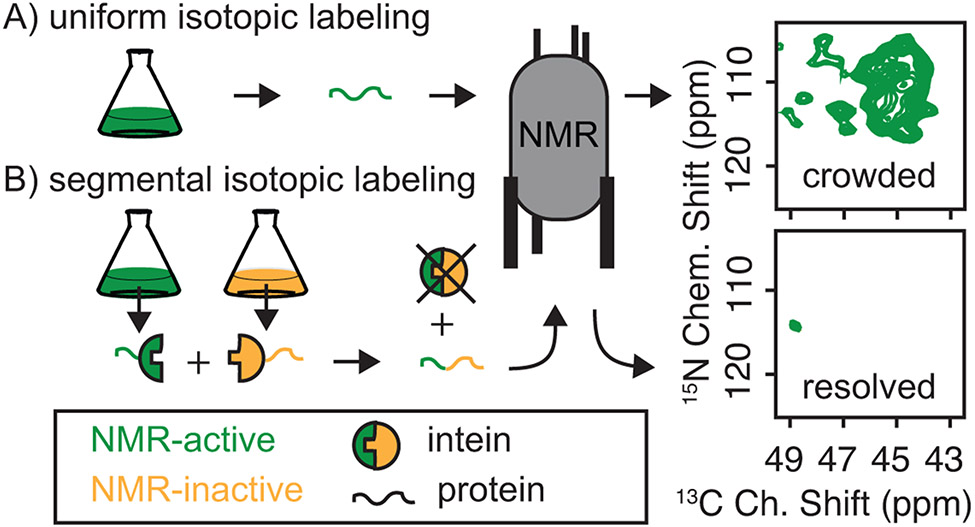
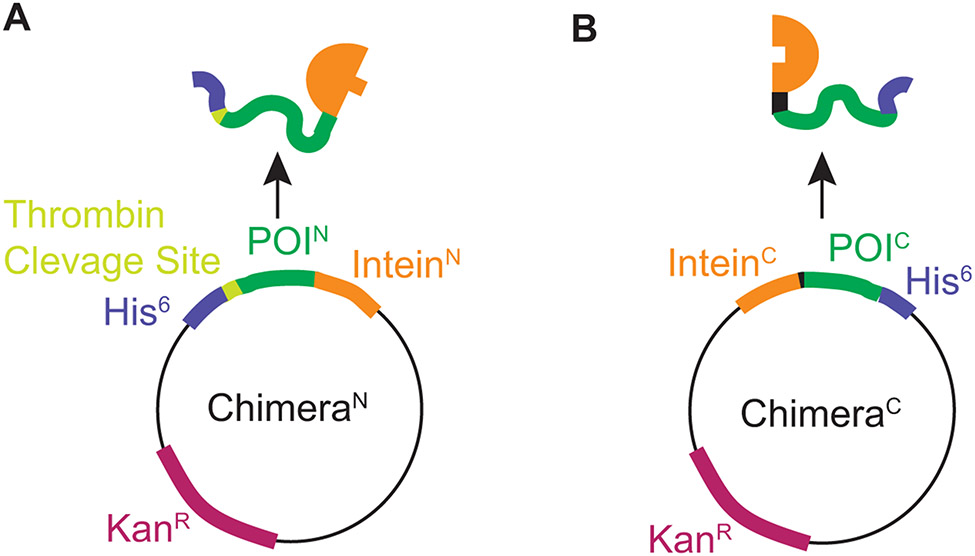
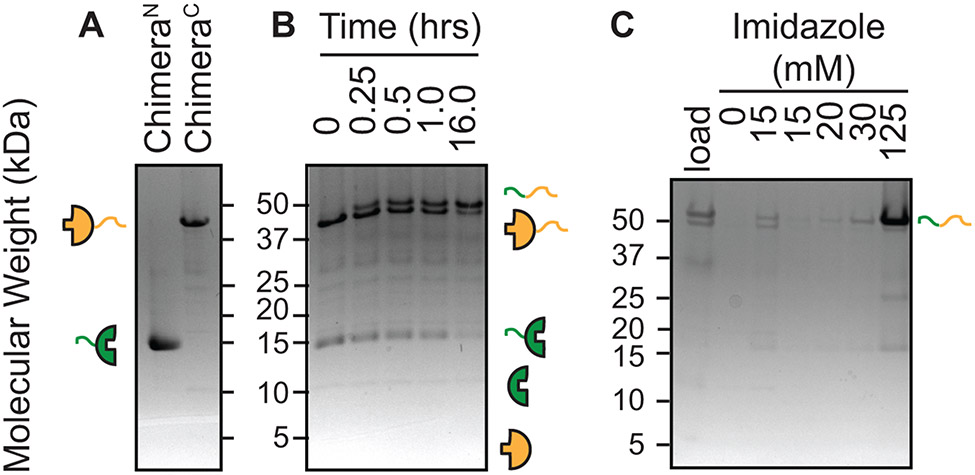
Structural investigations of biomolecules are typically confined to in vitro systems under extremely limited conditions. These investigations yield invaluable insights, but such experiments cannot capture important structural features imposed by cellular environments. Structural studies of proteins in their native contexts are not only possible using state-of-the-art sensitivity-enhanced (dynamic nuclear polarization, DNP) solid-state nuclear magnetic resonance (NMR) techniques, but these studies also demonstrate that the cellular context can and does have a dramatic influence on protein structure. In this chapter, we describe methods to prepare samples of isotopically labeled proteins at endogenous levels in cellular contexts alongside quality control methods to ensure that such samples accurately model important features of the cellular environment.
Keywords: Amyloids; Dynamic nuclear polarization; In situ structural biology; In-cell NMR; NMR; Prions; Sup35.
© 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- Abragam A (1983). The principles of nuclear magnetism. Clarendon Press.
-
- Bayro MJ, Debelouchina GT, Eddy MT, Birkett NR, MacPhee CE, Rosay M, et al. (2011). Intermolecular Structure Determination of Amyloid Fibrils with Magic-Angle Spinning and Dynamic Nuclear Polarization NMR. Journal of the American Chemical Society, 133(35), 13967–13974. 10.1021/ja203756x - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
