

Importin α Partitioning to the Plasma Membrane Regulates Intracellular Scaling
- PMID: 30639102
- PMCID: PMC6368448
- DOI: 10.1016/j.cell.2018.12.001
Importin α Partitioning to the Plasma Membrane Regulates Intracellular Scaling
Abstract
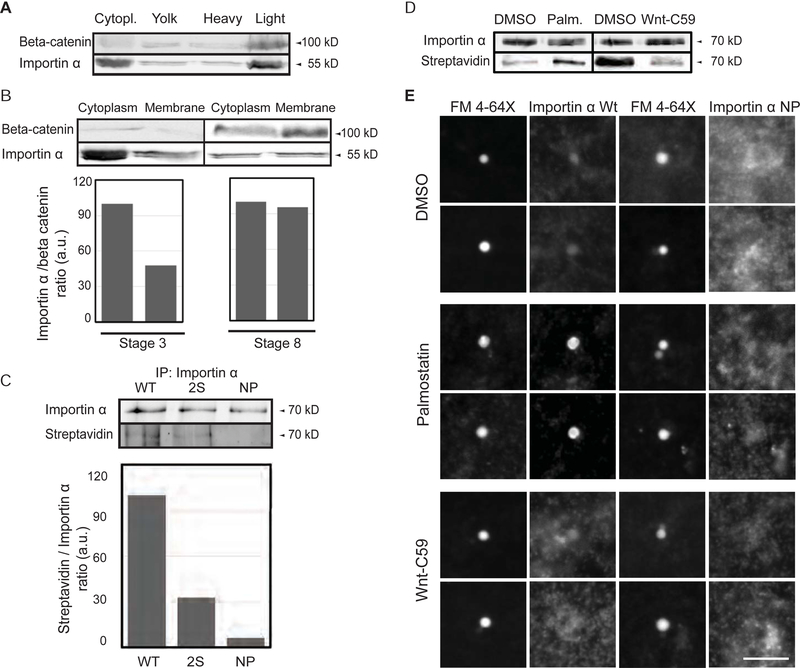
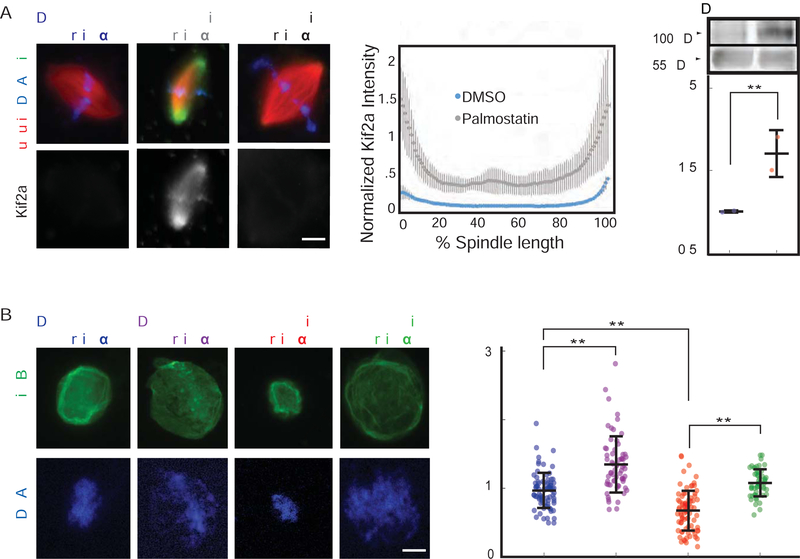
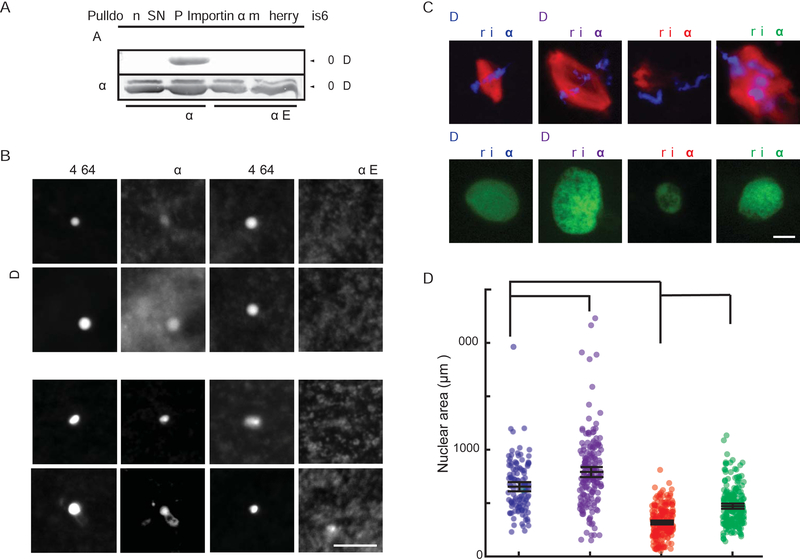
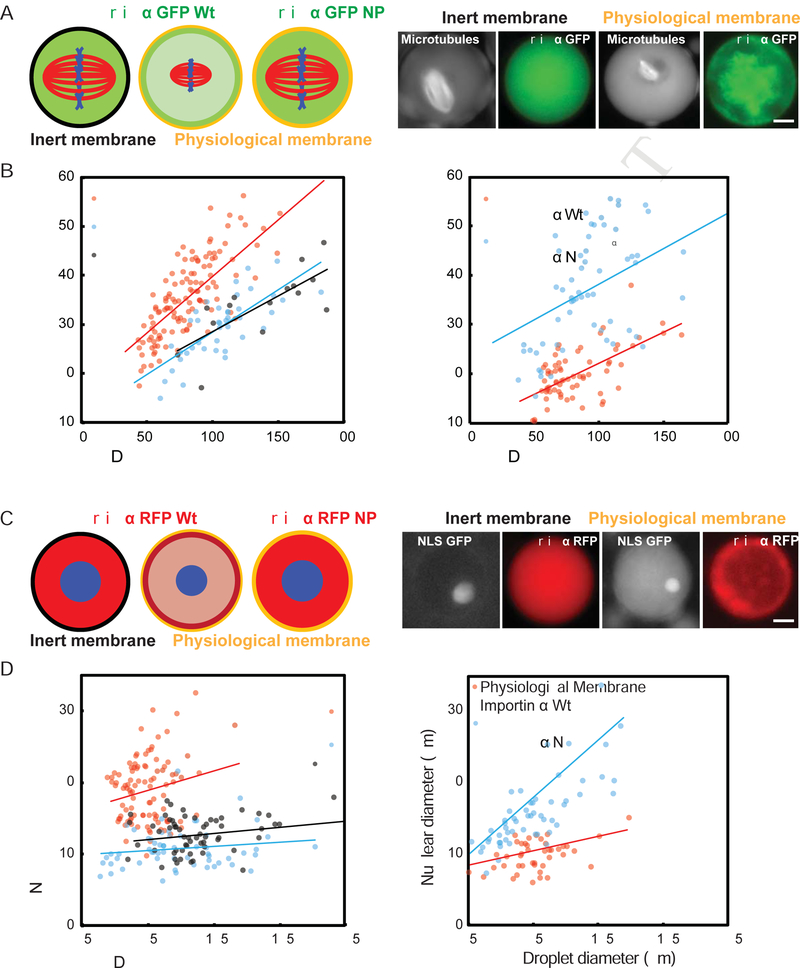
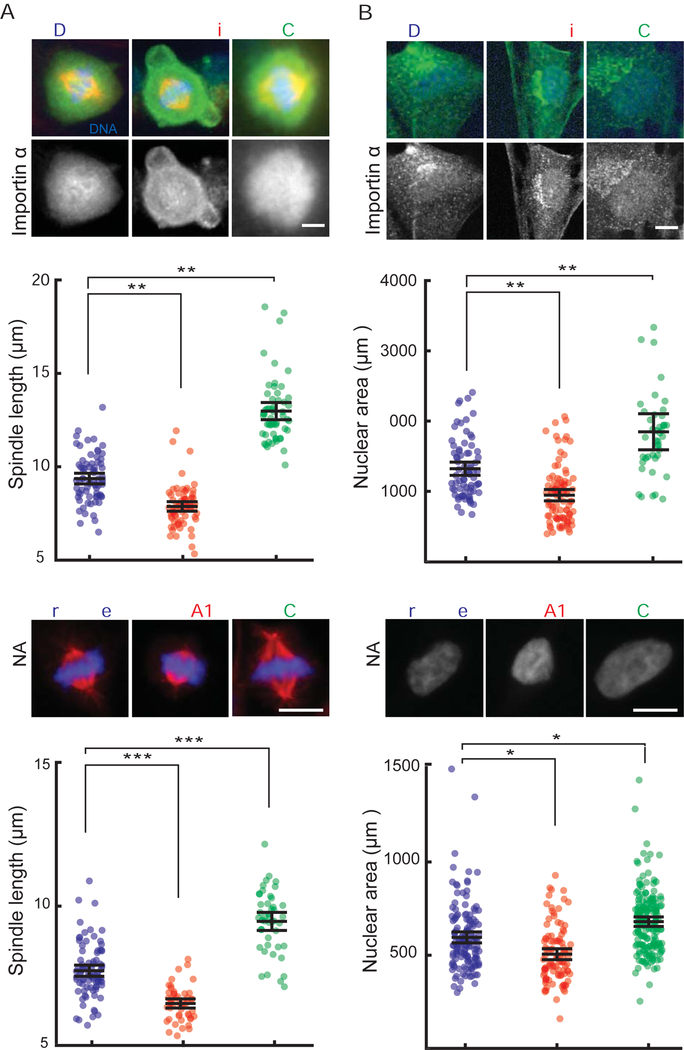
Early embryogenesis is accompanied by reductive cell divisions requiring that subcellular structures adapt to a range of cell sizes. The interphase nucleus and mitotic spindle scale with cell size through both physical and biochemical mechanisms, but control systems that coordinately scale intracellular structures are unknown. We show that the nuclear transport receptor importin α is modified by palmitoylation, which targets it to the plasma membrane and modulates its binding to nuclear localization signal (NLS)-containing proteins that regulate nuclear and spindle size in Xenopus egg extracts. Reconstitution of importin α targeting to the outer boundary of extract droplets mimicking cell-like compartments recapitulated scaling relationships observed during embryogenesis, which were altered by inhibitors that shift levels of importin α palmitoylation. Modulation of importin α palmitoylation in human cells similarly affected nuclear and spindle size. These experiments identify importin α as a conserved surface area-to-volume sensor that scales intracellular structures to cell size.
Keywords: KPNA2; Spindle scaling; casein kinase II; importin alpha; nuclear scaling; nuclear to cytoplasmic ratio; organelle scaling; palmitoylation.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures


Comment in
-
How cells keep scale.Nat Rev Mol Cell Biol. 2019 Mar;20(3):136. doi: 10.1038/s41580-019-0102-x. Nat Rev Mol Cell Biol. 2019. PMID: 30683908 No abstract available.
References
-
- Budde PP, Desai A, and Heald R (2005). Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods 38, 29–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
