

HIV-1 Reverse Transcriptase: A Metamorphic Protein with Three Stable States
- PMID: 30639227
- PMCID: PMC6686890
- DOI: 10.1016/j.str.2018.11.011
HIV-1 Reverse Transcriptase: A Metamorphic Protein with Three Stable States
Abstract
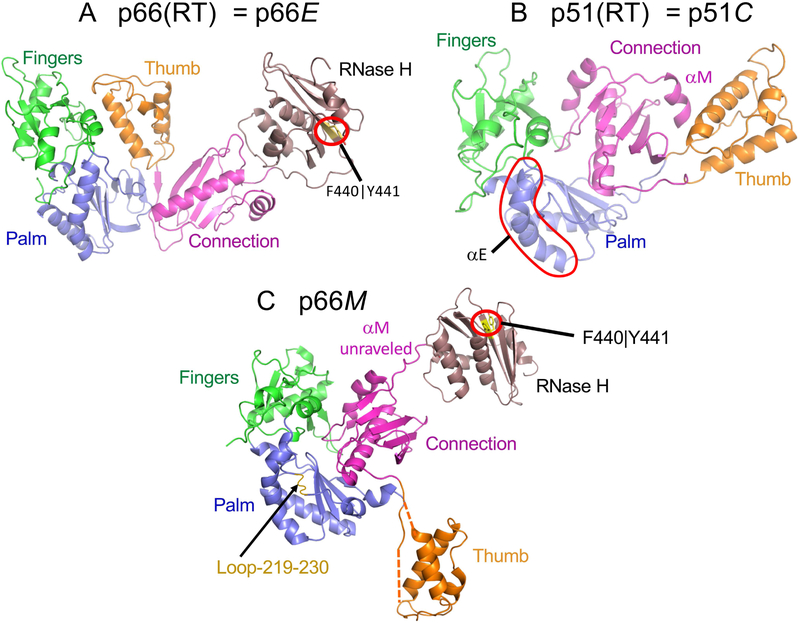
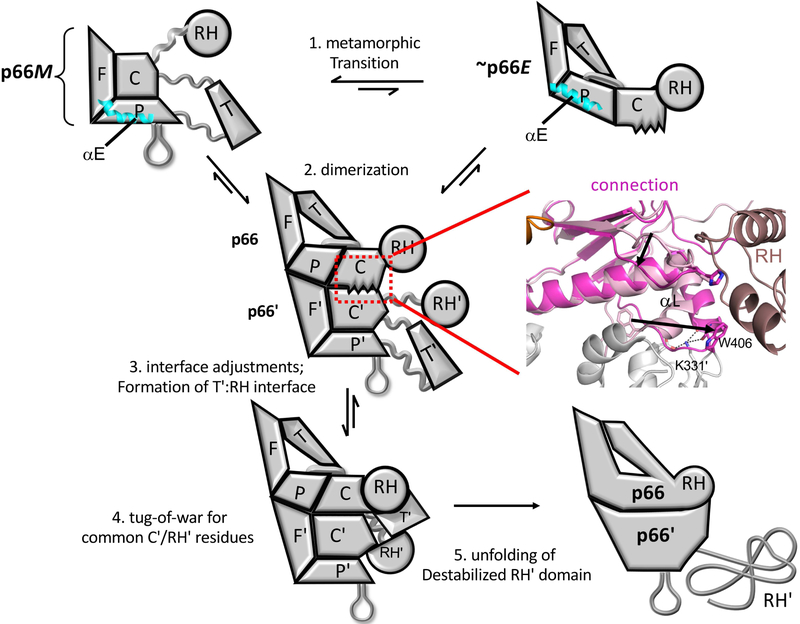
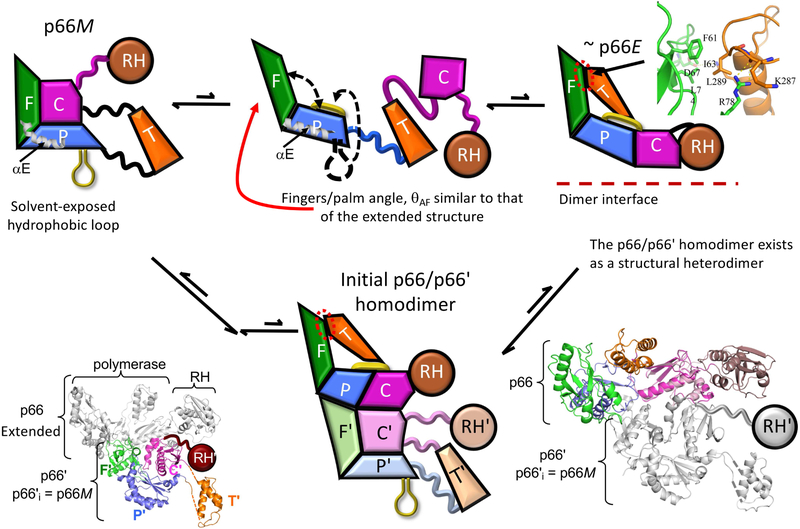
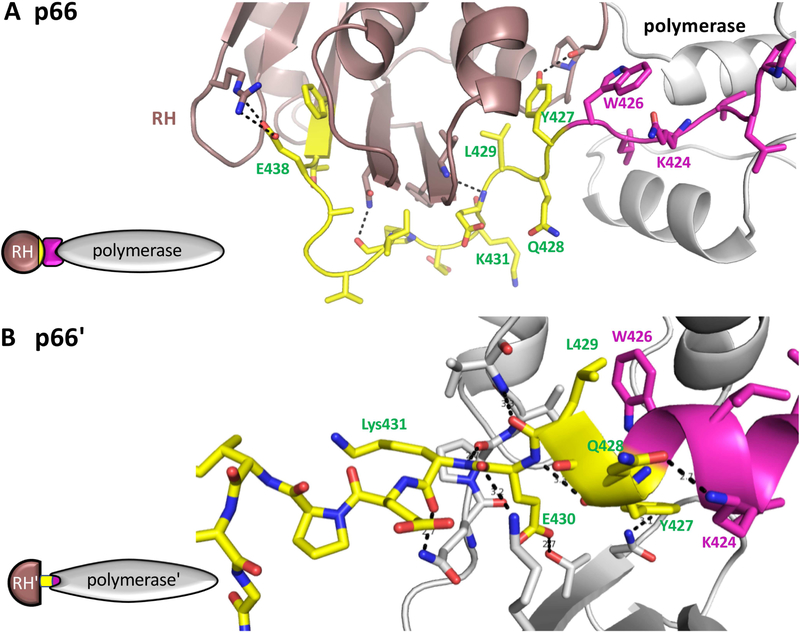
There has been a steadily increasing appreciation of the fact that the relationship between protein sequence and structure is often sufficiently ambiguous to allow a single sequence to adopt alternative, stable folds. Living organisms have been able to utilize such metamorphic proteins in remarkable and unanticipated ways. HIV-1 reverse transcriptase is among the earliest such proteins identified and remains a unique example in which a functional heterodimer contains two, alternatively folded polymerase domains. Structural characterization of the p66 precursor protein combined with NMR spectroscopic and molecular modeling studies have provided insights into the factors underlying the metamorphic transition and the subunit-specific programmed unfolding step required to expose the protease cleavage site within the ribonuclease H domain, supporting the conversion of the p66/p66' precursor into the mature p66/p51 heterodimer.
Keywords: HIV-1 reverse transcriptase; metamorphic protein; metamorphic transition; programmed unfolding; subunit-specific RH-domain unfolding.
Published by Elsevier Ltd.
Figures
References
-
- Becerra SP, Kumar A, Lewis MS, et al. (1991). Protein-protein interactions of HIV-1 reverse transcriptase: implication of central and C-terminal regions in subunit binding. Biochemistry-Us 30, 11707–11719. - PubMed
-
- Choi H, Kim S, Mukhopadhyay P, et al. (2001). Structural basis of the redox switch in the OxyR transcription factor. Cell 105, 103–113. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
