

Hepatocyte-specific lysosomal acid lipase deficiency protects mice from diet-induced obesity but promotes hepatic inflammation
- PMID: 30639734
- PMCID: PMC6372077
- DOI: 10.1016/j.bbalip.2019.01.007
Hepatocyte-specific lysosomal acid lipase deficiency protects mice from diet-induced obesity but promotes hepatic inflammation
Abstract
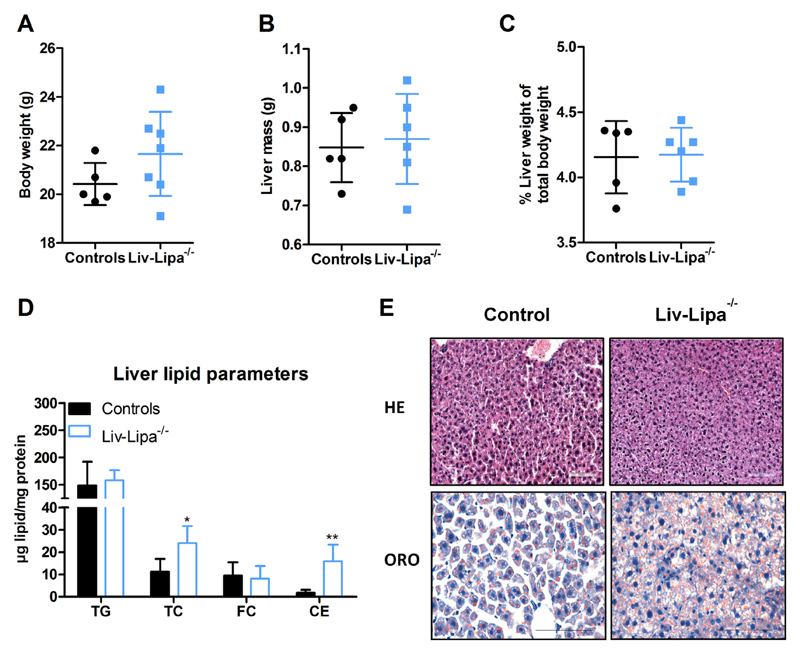
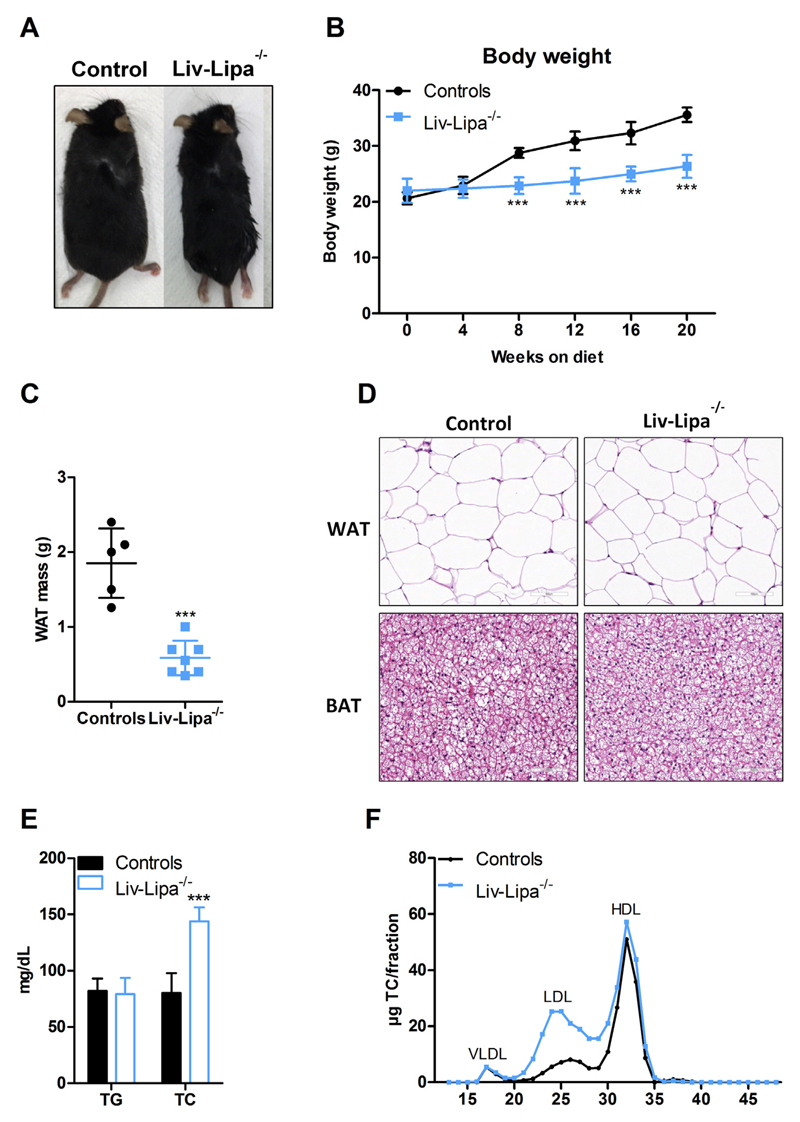
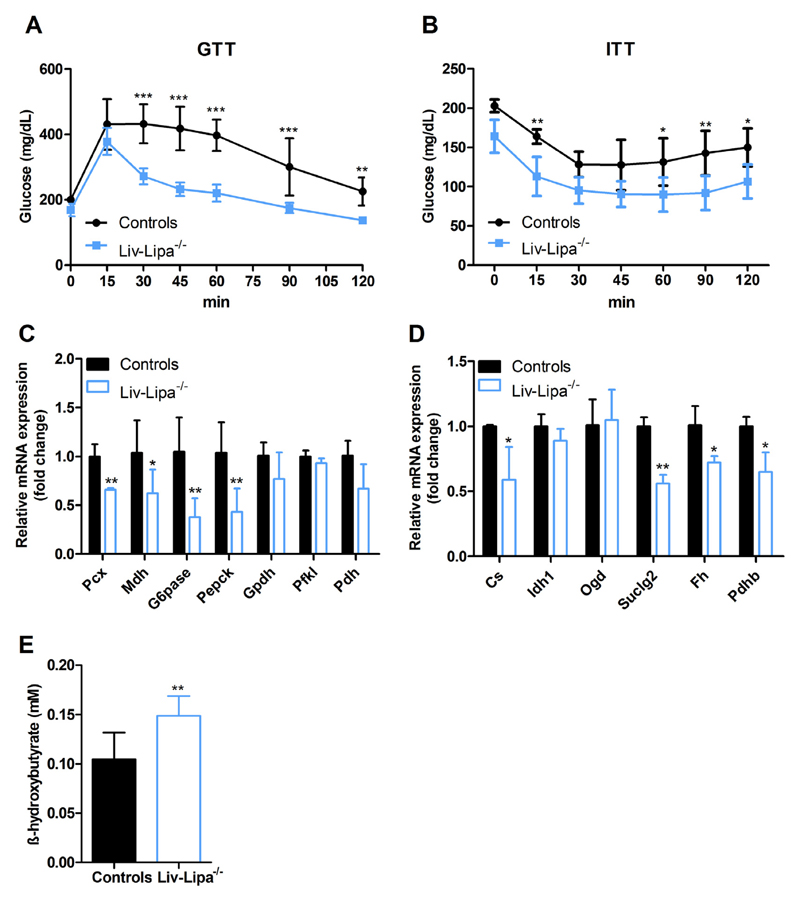
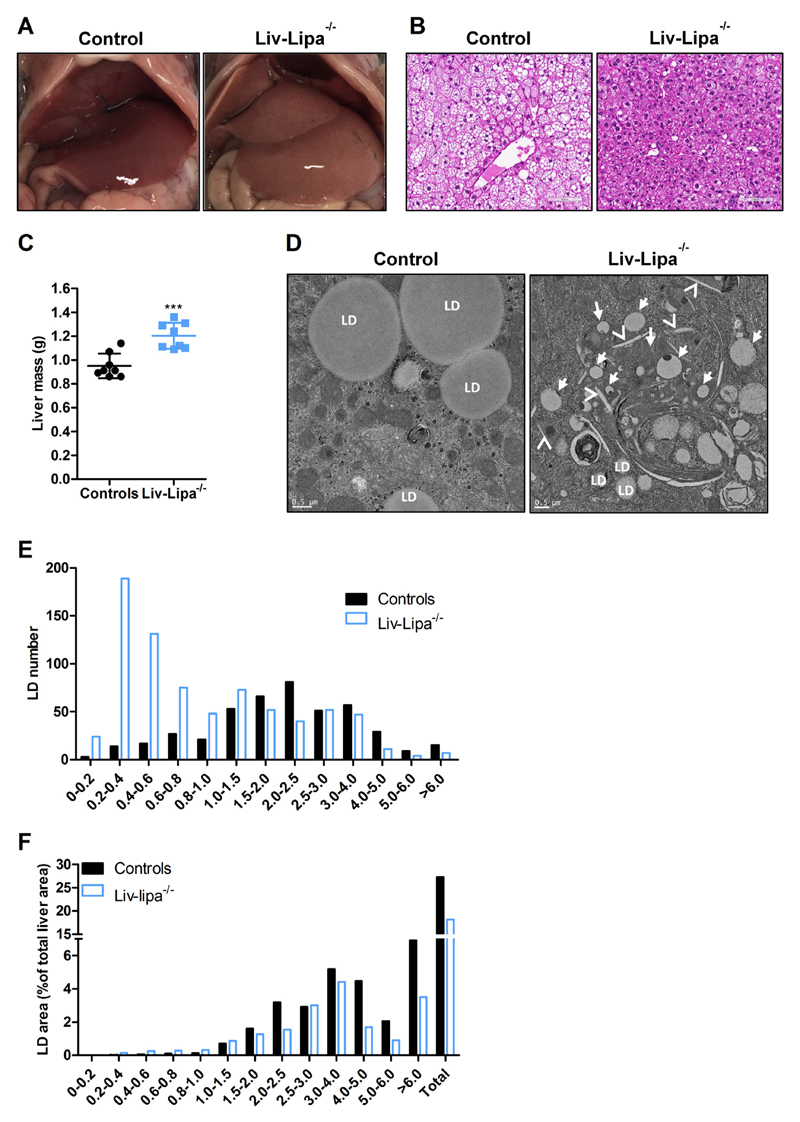
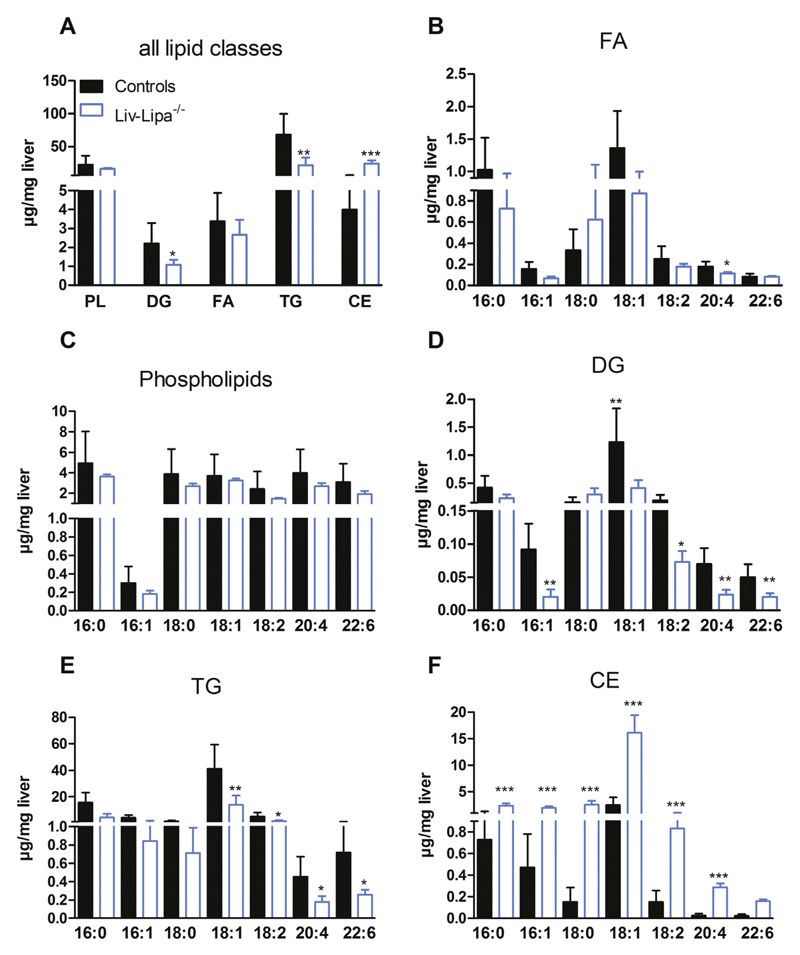
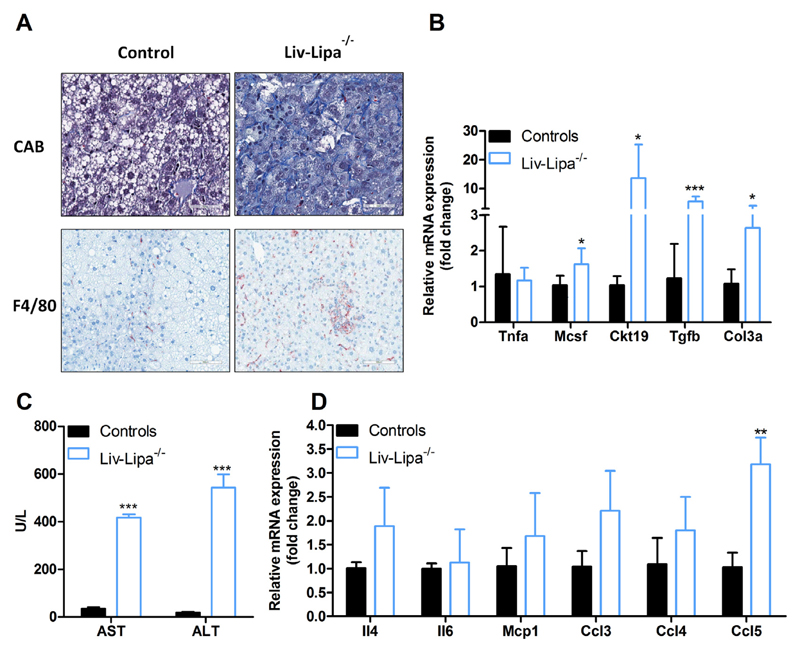
Lysosomal acid lipase (LAL) hydrolyzes cholesteryl esters (CE) and triglycerides (TG) to generate fatty acids (FA) and cholesterol. LAL deficiency (LAL-D) in both humans and mice leads to hepatomegaly, hypercholesterolemia, and shortened life span. Despite its essential role in lysosomal neutral lipid catabolism, the cell type-specific contribution of LAL to disease progression is still elusive. To investigate the role of LAL in the liver in more detail and to exclude the contribution of LAL in macrophages, we generated hepatocyte-specific LAL-deficient mice (Liv-Lipa-/-) and fed them either chow or high fat/high cholesterol diets (HF/HCD). Comparable to systemic LAL-D, Liv-Lipa-/- mice were resistant to diet-induced obesity independent of food intake, movement, and energy expenditure. Reduced body weight gain was mainly due to reduced white adipose tissue depots. Furthermore, Liv-Lipa-/- mice exhibited improved glucose clearance during glucose and insulin tolerance tests compared to control mice. Analysis of hepatic lipid content revealed a massive reduction of TG, whereas CE concentrations were markedly increased, leading to CE crystal formation in the livers of Liv-Lipa-/- mice. Elevated plasma transaminase activities, increased pro-inflammatory cytokines and chemokines as well as hepatic macrophage infiltration indicated liver inflammation. Our data provide evidence that hepatocyte-specific LAL deficiency is sufficient to alter whole-body lipid and energy homeostasis in mice. We conclude that hepatic LAL plays a pivotal role by preventing liver damage and maintaining lipid and energy homeostasis, especially during high lipid availability.
Keywords: Cholesteryl ester storage disease; Fibrosis; LAL-D; Lipid; Liver damage; Wolman disease.
Copyright © 2019 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare that there is no conflict of interest associated with this manuscript.
Figures

References
-
- Sheriff S, Du H, Grabowski GA. Characterization of lysosomal acid lipase by site-directed mutagenesis and heterologous expression. J Biol Chem. 1995;270:27766–27772. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
