

The role of microtubules and the dynein/dynactin motor complex of host cells in the biogenesis of the Coxiella burnetii-containing vacuole
- PMID: 30640917
- PMCID: PMC6331085
- DOI: 10.1371/journal.pone.0209820
The role of microtubules and the dynein/dynactin motor complex of host cells in the biogenesis of the Coxiella burnetii-containing vacuole
Abstract
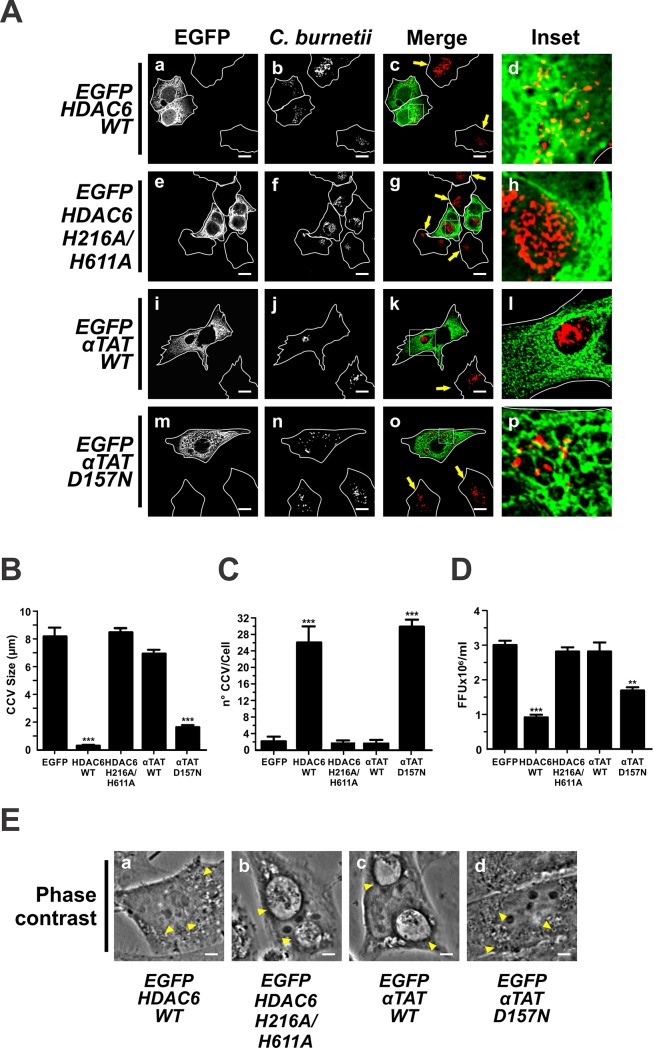
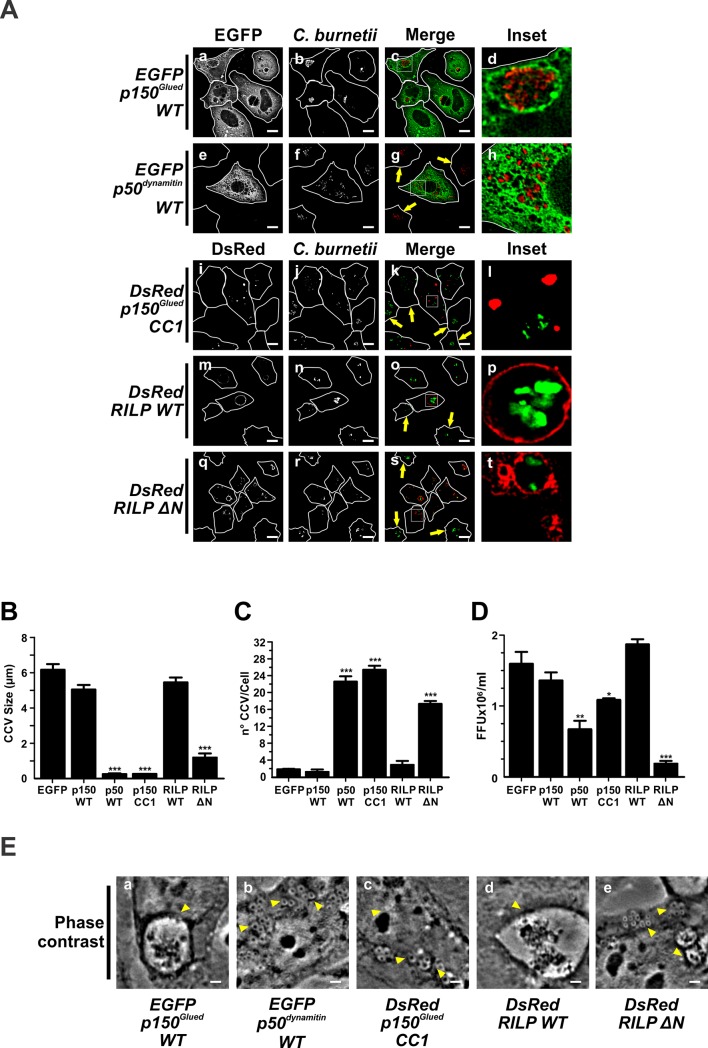
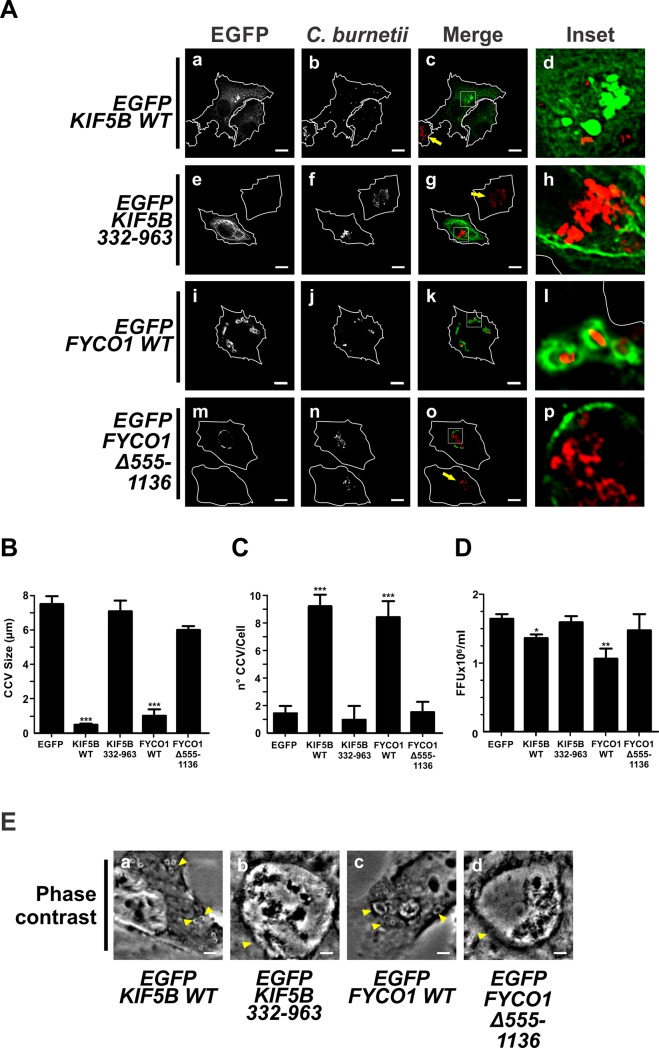
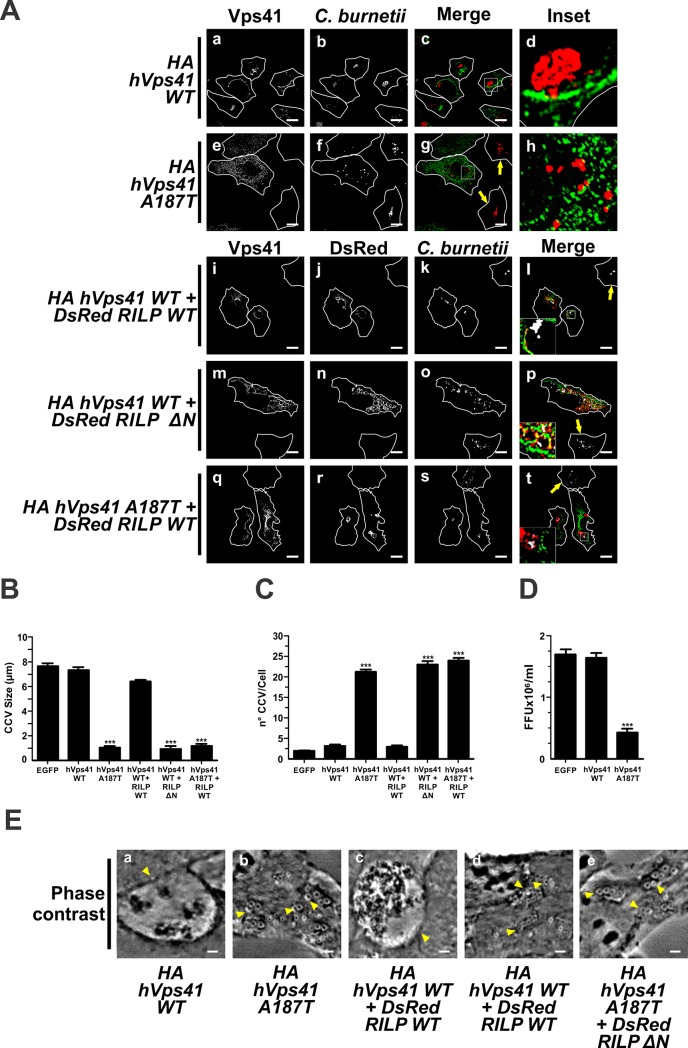
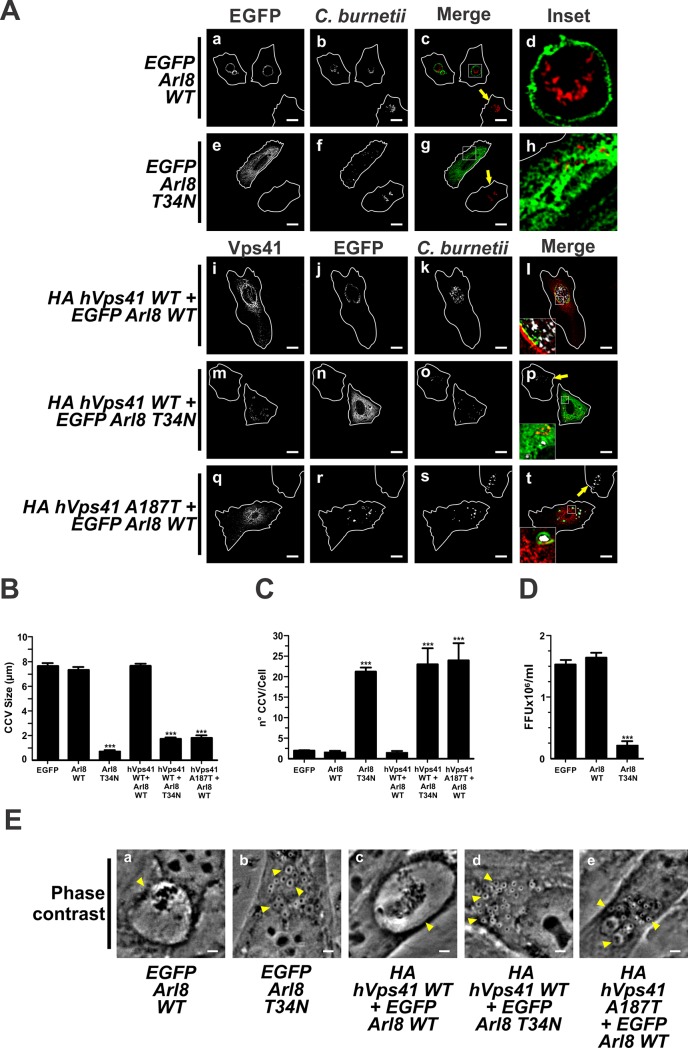
Microtubules (Mts) are dynamic cytoskeleton structures that play a key role in vesicular transport. The Mts-mediated transport depends on motor proteins named kinesins and the dynein/dynactin motor complex. The Rab7 adapter protein FYCO1 controls the anterograde transport of the endocytic compartments through the interaction with the kinesin KIF5. Rab7 and its partner RILP induce the recruitment of dynein/dynactin to late endosomes regulating its retrograde transport to the perinuclear area to fuse with lysosomes. The late endosomal-lysosomal fusion is regulated by the HOPS complex through its interaction with RILP and the GTPase Arl8. Coxiella burnetii (Cb), the causative agent of Q fever, is an obligate intracellular pathogen, which generates a large compartment with autophagolysosomal characteristics named Cb-containing vacuole (CCV). The CCV forms through homotypic fusion between small non-replicative CCVs (nrCCV) and through heterotypic fusion with other compartments, such as endosomes and lysosomes. In this work, we characterise the role of Mts, motor proteins, RILP/Rab7 and Arl8 on the CCV biogenesis. The formation of the CCV was affected when either the dynamics and/or the acetylation state of Mts were modified. Similarly, the overexpression of the dynactin subunit non-functional mutants p150Glued and RILP led to the formation of small nrCCVs. This phenomenon is not observed in cells overexpressing WT proteins, the motor KIF5 or its interacting protein FYCO1. The formation of the CCV was normal in infected cells that overexpressed Arl8 alone or together with hVps41 (a HOPS subunit) or in cells co-overexpressing hVps41 and RILP. The dominant negative mutant of Arl8 and the non-functional hVps41 inhibited the formation of the CCV. When the formation of CCV was affected, the bacterial multiplication diminished. Our results suggest that nrCCVs recruit the molecular machinery that regulate the Mts-dependent retrograde transport, Rab7/RILP and the dynein/dynactin system, as well as the tethering processes such as HOPS complex and Arl8 to finally originate the CCV where C. burnetii multiplies.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
