

Gene therapy targeting SARM1 blocks pathological axon degeneration in mice
- PMID: 30642945
- PMCID: PMC6363435
- DOI: 10.1084/jem.20181040
Gene therapy targeting SARM1 blocks pathological axon degeneration in mice
Abstract
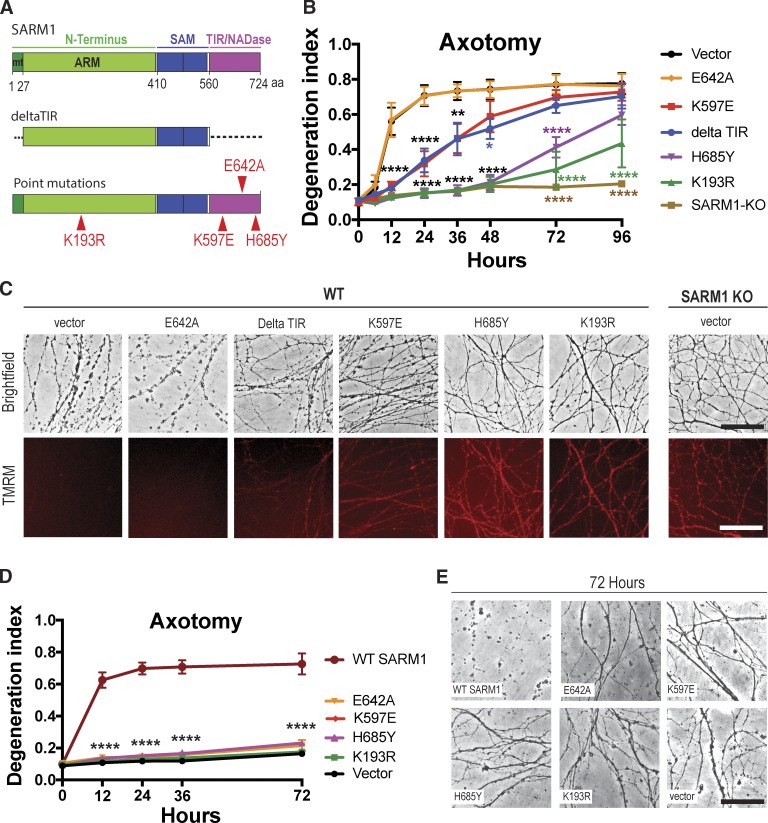
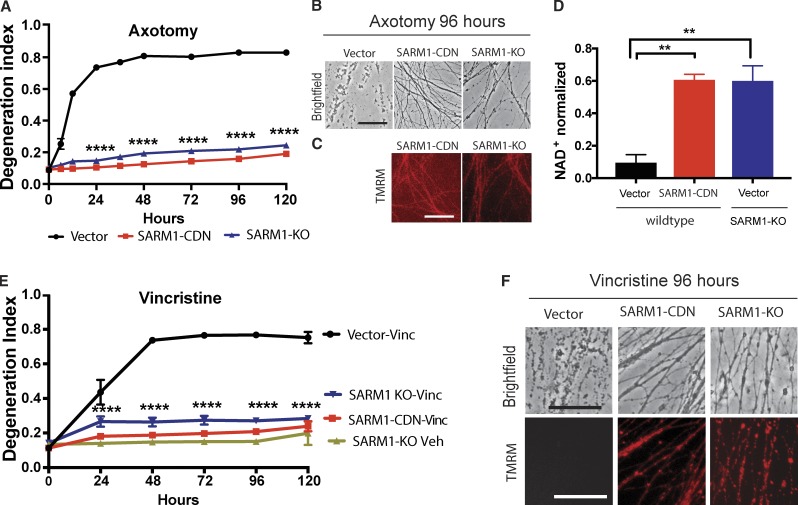
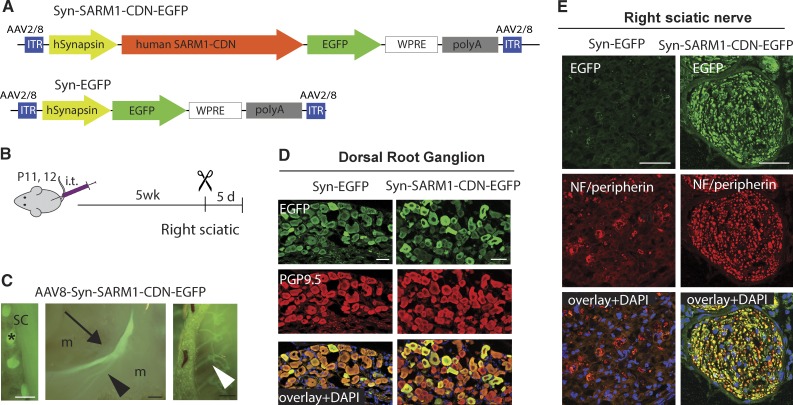
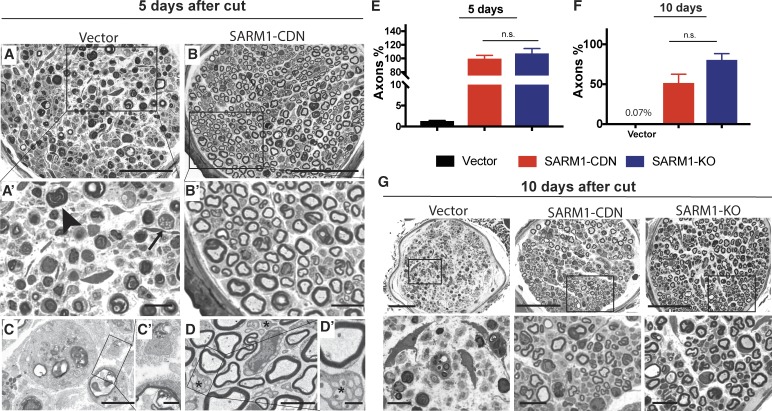
Axonal degeneration (AxD) following nerve injury, chemotherapy, and in several neurological disorders is an active process driven by SARM1, an injury-activated NADase. Axons of SARM1-null mice exhibit greatly delayed AxD after transection and in models of neurological disease, suggesting that inhibiting SARM1 is a promising strategy to reduce pathological AxD. Unfortunately, no drugs exist to target SARM1. We, therefore, developed SARM1 dominant-negatives that potently block AxD in cellular models of axotomy and neuropathy. To assess efficacy in vivo, we used adeno-associated virus-mediated expression of the most potent SARM1 dominant-negative and nerve transection as a model of severe AxD. While axons of vehicle-treated mice degenerate rapidly, axons of mice expressing SARM1 dominant-negative can remain intact for >10 d after transection, similar to the protection observed in SARM1-null mice. We thus developed a novel in vivo gene therapeutic to block pathological axon degeneration by inhibiting SARM1, an approach that may be applied clinically to treat manifold neurodegenerative diseases characterized by axon loss.
© 2019 Geisler et al.
Figures
References
-
- Caminiti S.P., Presotto L., Baroncini D., Garibotto V., Moresco R.M., Gianolli L., Volonté M.A., Antonini A., and Perani D.. 2017. Axonal damage and loss of connectivity in nigrostriatal and mesolimbic dopamine pathways in early Parkinson’s disease. Neuroimage Clin. 14:734–740. 10.1016/j.nicl.2017.03.011 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
