

A macrophage-based screen identifies antibacterial compounds selective for intracellular Salmonella Typhimurium
- PMID: 30643129
- PMCID: PMC6331611
- DOI: 10.1038/s41467-018-08190-x
A macrophage-based screen identifies antibacterial compounds selective for intracellular Salmonella Typhimurium
Abstract
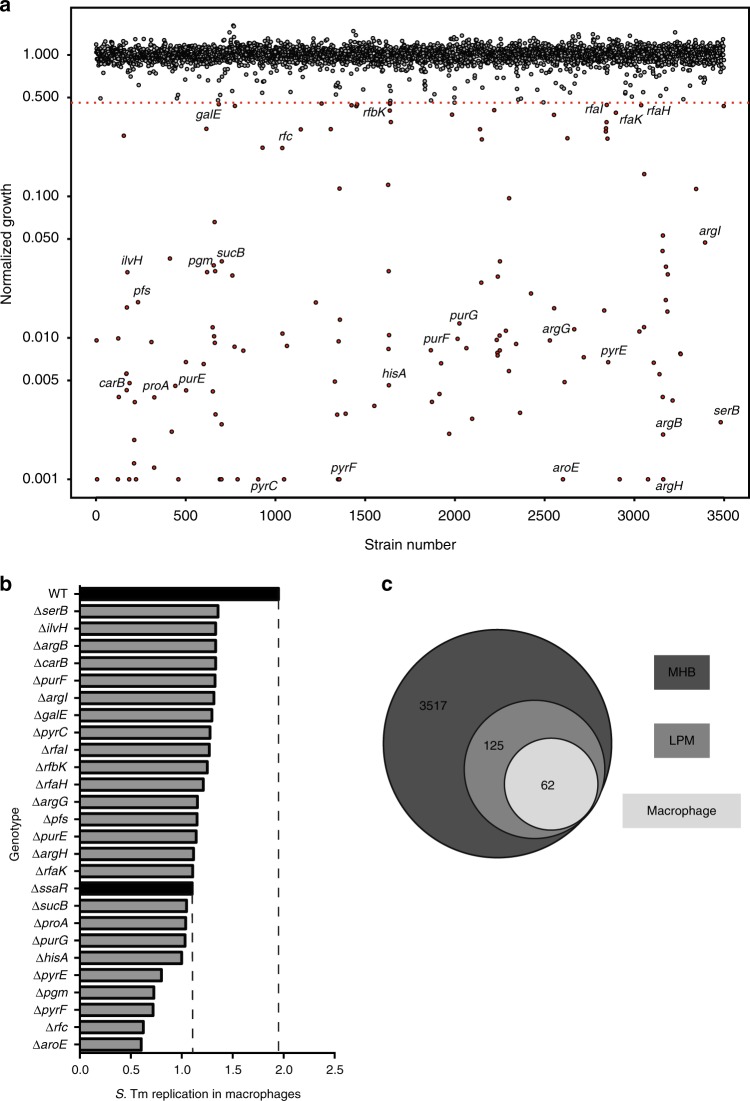
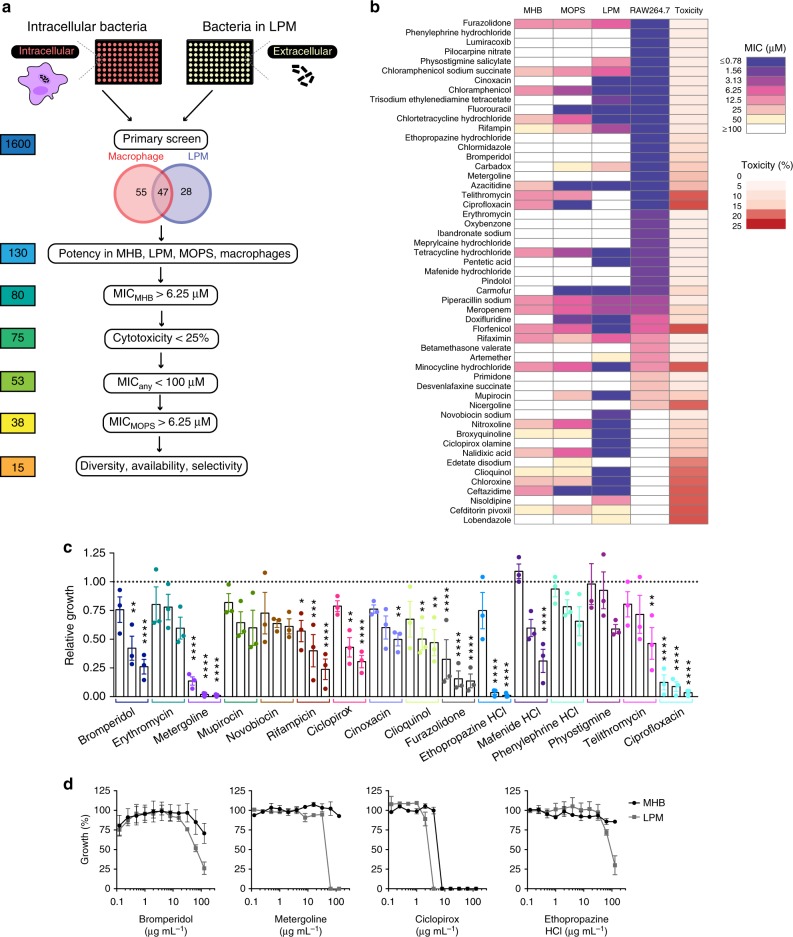
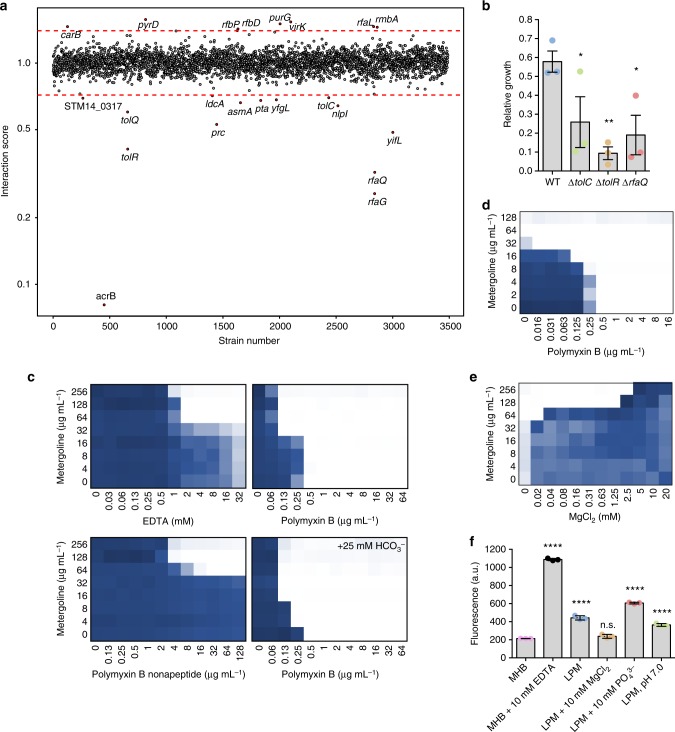
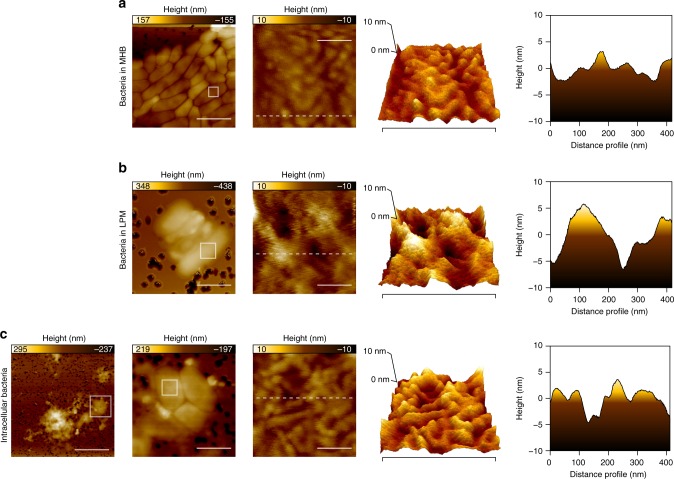
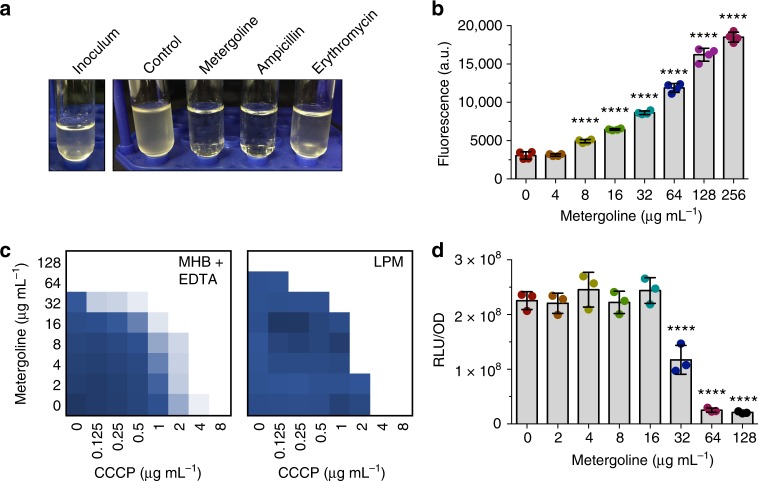
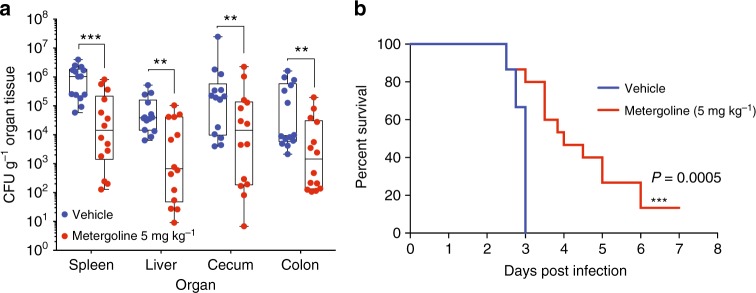
Salmonella Typhimurium (S. Tm) establishes systemic infection in susceptible hosts by evading the innate immune response and replicating within host phagocytes. Here, we sought to identify inhibitors of intracellular S. Tm replication by conducting parallel chemical screens against S. Tm growing in macrophage-mimicking media and within macrophages. We identify several compounds that inhibit Salmonella growth in the intracellular environment and in acidic, ion-limited media. We report on the antimicrobial activity of the psychoactive drug metergoline, which is specific against intracellular S. Tm. Screening an S. Tm deletion library in the presence of metergoline reveals hypersensitization of outer membrane mutants to metergoline activity. Metergoline disrupts the proton motive force at the bacterial cytoplasmic membrane and extends animal survival during a systemic S. Tm infection. This work highlights the predictive nature of intracellular screens for in vivo efficacy, and identifies metergoline as a novel antimicrobial active against Salmonella.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
