

Metabolome and transcriptome-wide effects of the carbon storage regulator A in enteropathogenic Escherichia coli
- PMID: 30644424
- PMCID: PMC6333774
- DOI: 10.1038/s41598-018-36932-w
Metabolome and transcriptome-wide effects of the carbon storage regulator A in enteropathogenic Escherichia coli
Abstract
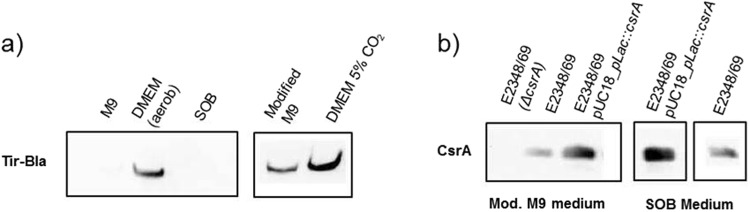
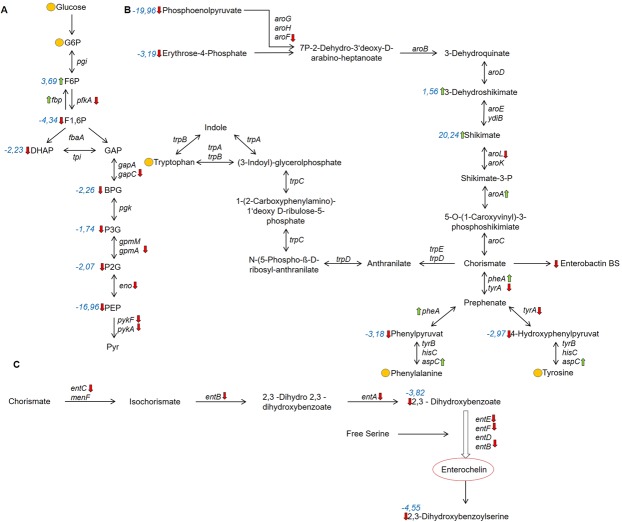
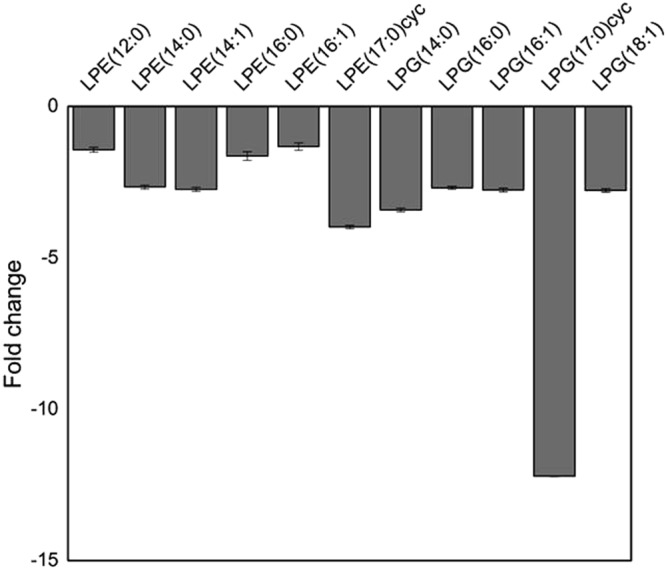
The carbon storage regulator A (CsrA) is a conserved global regulatory system known to control central carbon pathways, biofilm formation, motility, and pathogenicity. The aim of this study was to characterize changes in major metabolic pathways induced by CsrA in human enteropathogenic Escherichia coli (EPEC) grown under virulence factor-inducing conditions. For this purpose, the metabolomes and transcriptomes of EPEC and an isogenic ∆csrA mutant derivative were analyzed by untargeted mass spectrometry and RNA sequencing, respectively. Of the 159 metabolites identified from untargeted GC/MS and LC/MS data, 97 were significantly (fold change ≥ 1.5; corrected p-value ≤ 0.05) regulated between the knockout and the wildtype strain. A lack of csrA led to an accumulation of fructose-6-phosphate (F6P) and glycogen synthesis pathway products, whereas metabolites in lower glycolysis and the citric acid cycle were downregulated. Associated pathways from the citric acid cycle like aromatic amino acid and siderophore biosynthesis were also negatively influenced. The nucleoside salvage pathways were featured by an accumulation of nucleosides and nucleobases, and a downregulation of nucleotides. In addition, a pronounced downregulation of lyso-lipid metabolites was observed. A drastic change in the morphology in the form of vesicle-like structures of the ∆csrA knockout strain was visible by electron microscopy. Colanic acid synthesis genes were strongly (up to 50 fold) upregulated, and the abundance of colanic acid was 3 fold increased according to a colorimetric assay. The findings expand the scope of pathways affected by the csrA regulon and emphasize its importance as a global regulator.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Rothbaum R, McAdams AJ, Giannella R, Partin JC. A clinicopathologic study of enterocyte-adherent Escherichia coli: a cause of protracted diarrhea in infants. Gastroenterology. 1982;83:441–454. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
