

Fucosylation Enhances the Efficacy of Adoptively Transferred Antigen-Specific Cytotoxic T Lymphocytes
- PMID: 30647079
- PMCID: PMC6467811
- DOI: 10.1158/1078-0432.CCR-18-1527
Fucosylation Enhances the Efficacy of Adoptively Transferred Antigen-Specific Cytotoxic T Lymphocytes
Abstract
Purpose: Inefficient homing of adoptively transferred cytotoxic T lymphocytes (CTLs) to tumors is a major limitation to the efficacy of adoptive cellular therapy (ACT) for cancer. However, through fucosylation, a process whereby fucosyltransferases (FT) add fucose groups to cell surface glycoproteins, this challenge may be overcome. Endogenously fucosylated CTLs and ex vivo fucosylated cord blood stem cells and regulatory T cells were shown to preferentially home to inflamed tissues and marrow. Here, we show a novel approach to enhance CTL homing to leukemic marrow and tumor tissue.
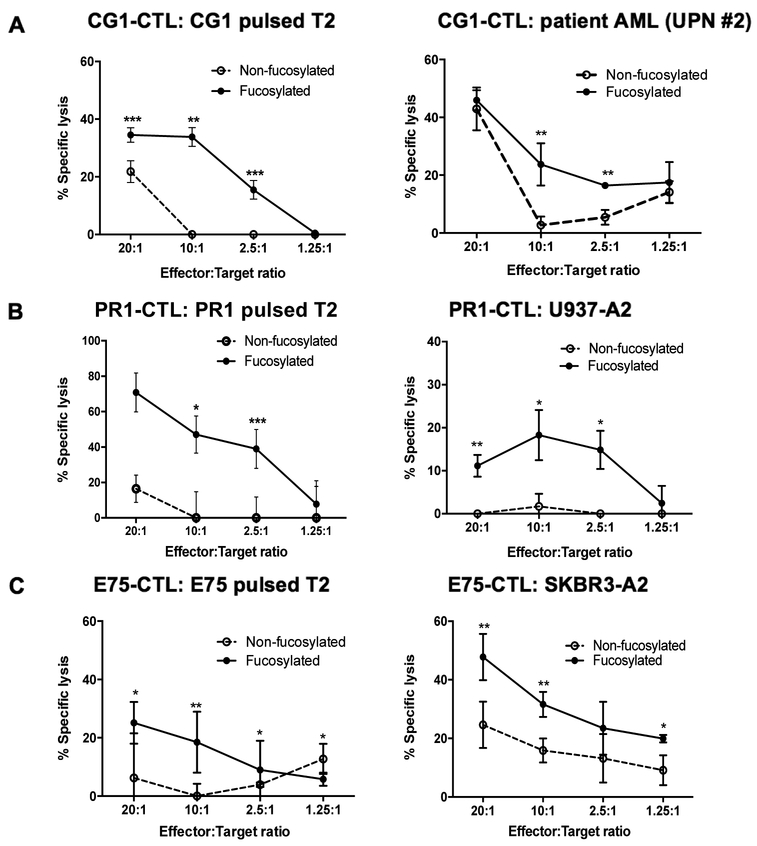
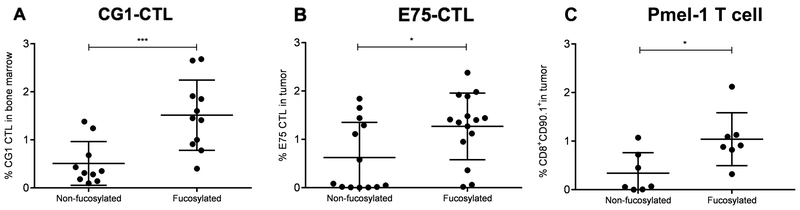
Experimental design: Using the enzyme FT-VII, we fucosylated CTLs that target the HLA-A2-restricted leukemia antigens CG1 and PR1, the HER2-derived breast cancer antigen E75, and the melanoma antigen gp-100. We performed in vitro homing assays to study the effects of fucosylation on CTL homing and target killing. We used in vivo mouse models to demonstrate the effects of ex vivo fucosylation on CTL antitumor activities against leukemia, breast cancer, and melanoma.
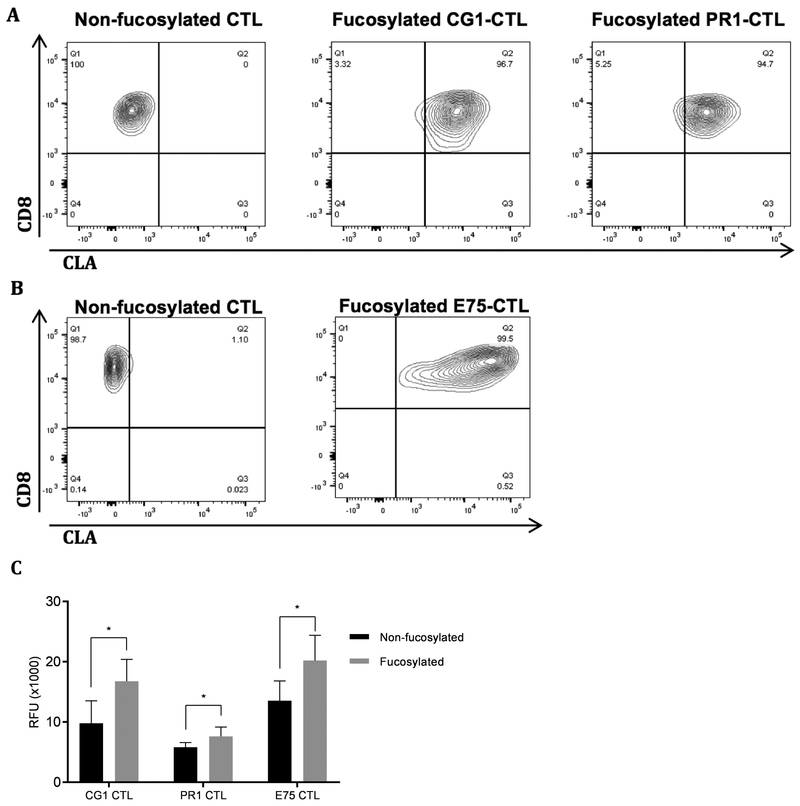
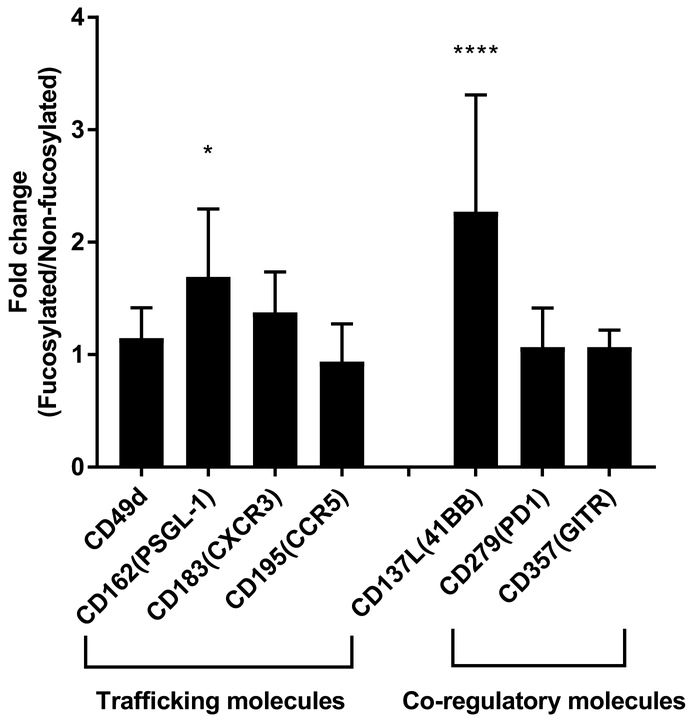
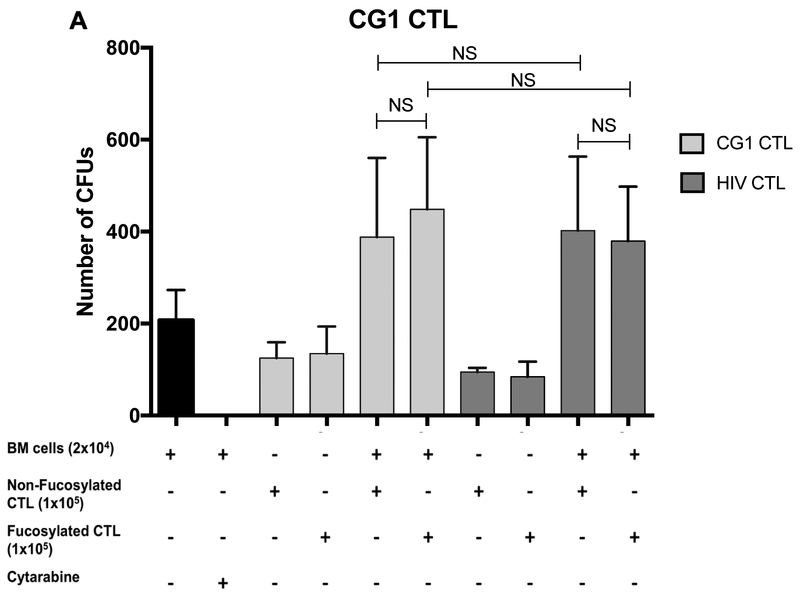
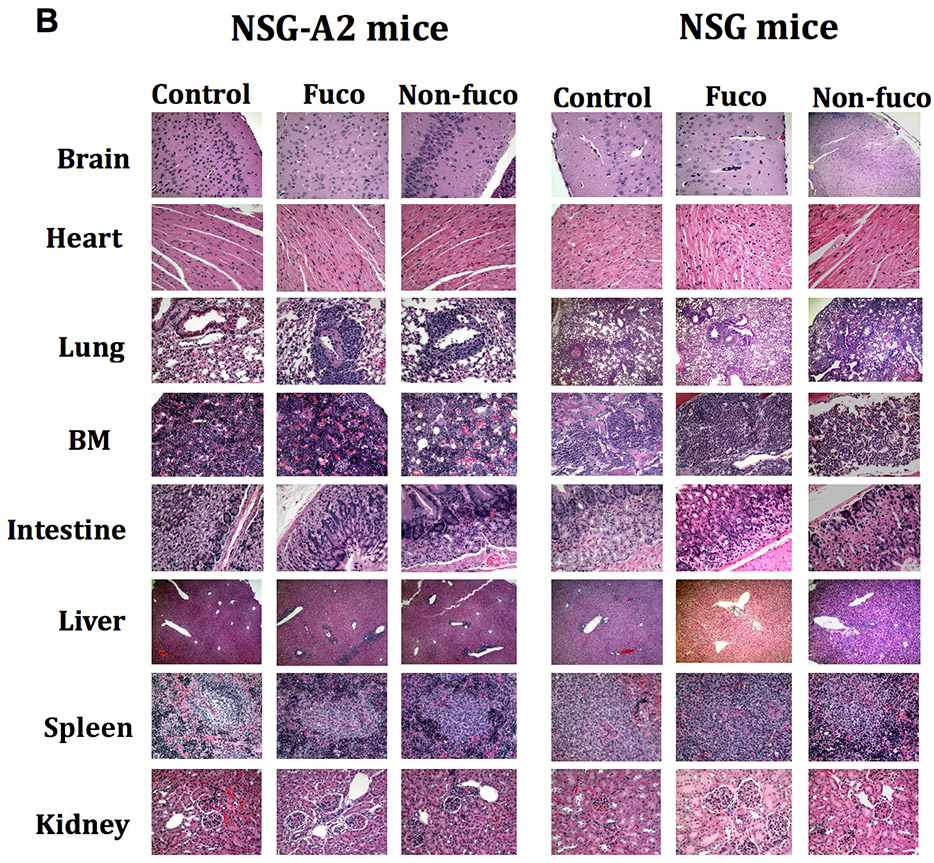
Results: Our data show that fucosylation increases in vitro homing and cytotoxicity of antigen-specific CTLs. Furthermore, fucosylation enhances in vivo CTL homing to leukemic bone marrow, breast cancer, and melanoma tissue in NOD/SCID gamma (NSG) and immunocompetent mice, ultimately boosting the antitumor activity of the antigen-specific CTLs. Importantly, our work demonstrates that fucosylation does not interfere with CTL specificity.
Conclusions: Together, our data establish ex vivo CTL fucosylation as a novel approach to improving the efficacy of ACT, which may be of great value for the future of ACT for cancer.
©2019 American Association for Cancer Research.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
