

On the Mechanism of Hyperthermia-Induced BRCA2 Protein Degradation
- PMID: 30650591
- PMCID: PMC6356811
- DOI: 10.3390/cancers11010097
On the Mechanism of Hyperthermia-Induced BRCA2 Protein Degradation
Abstract
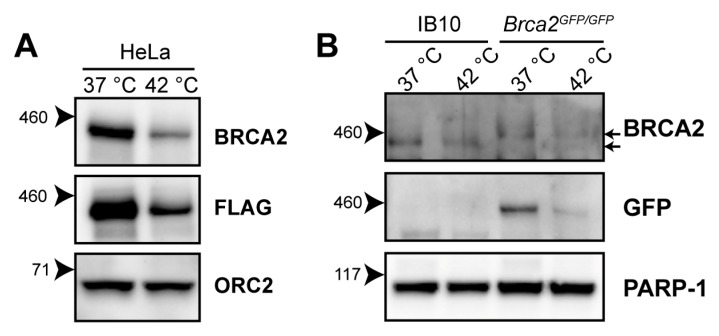
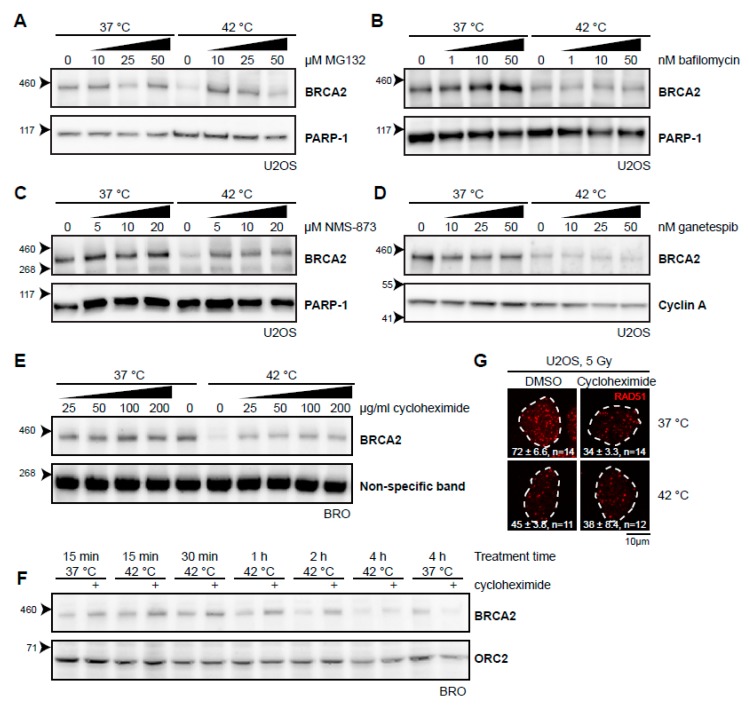
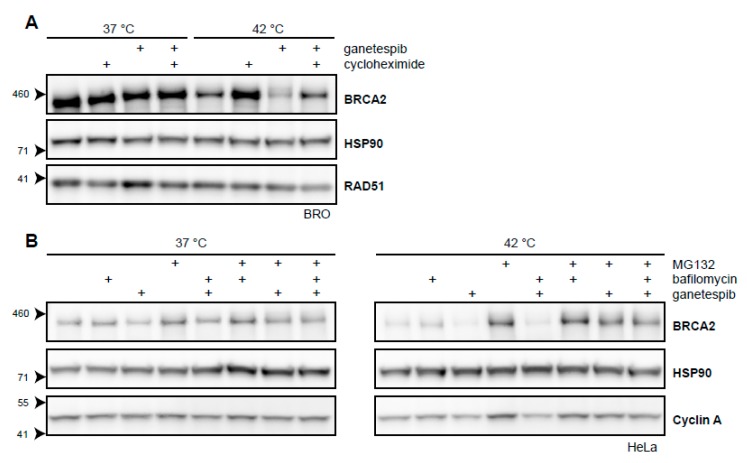
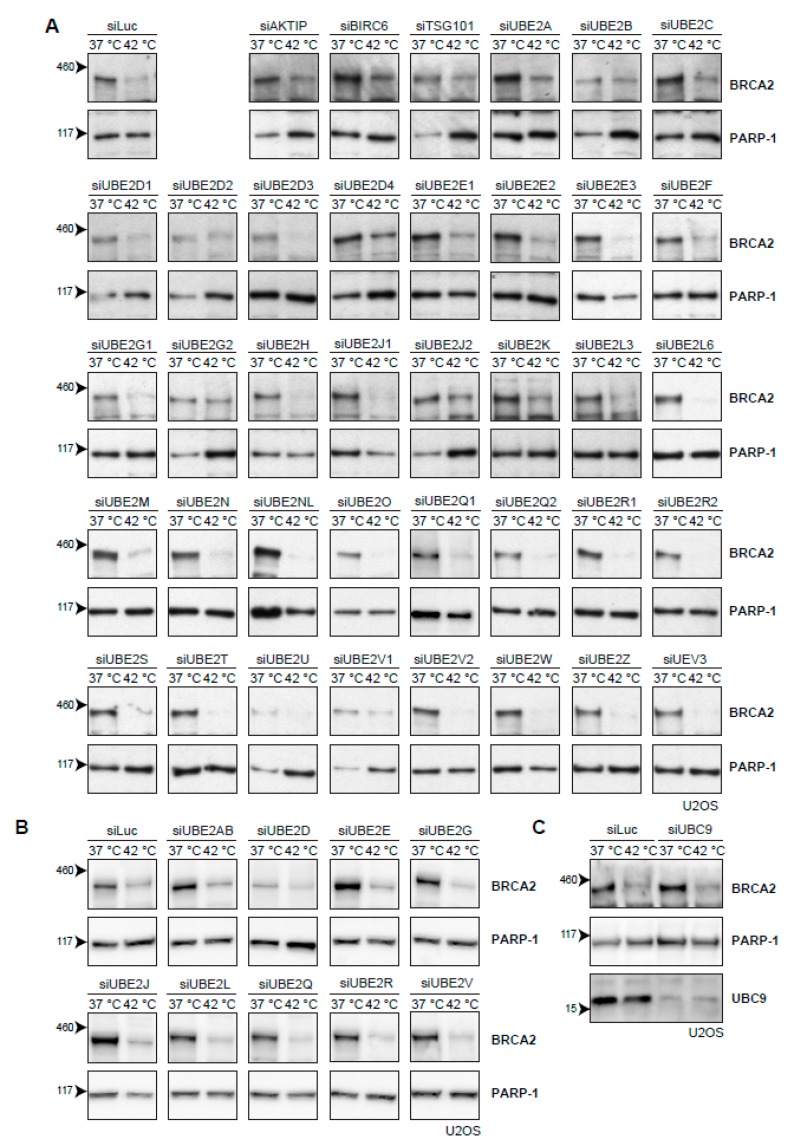
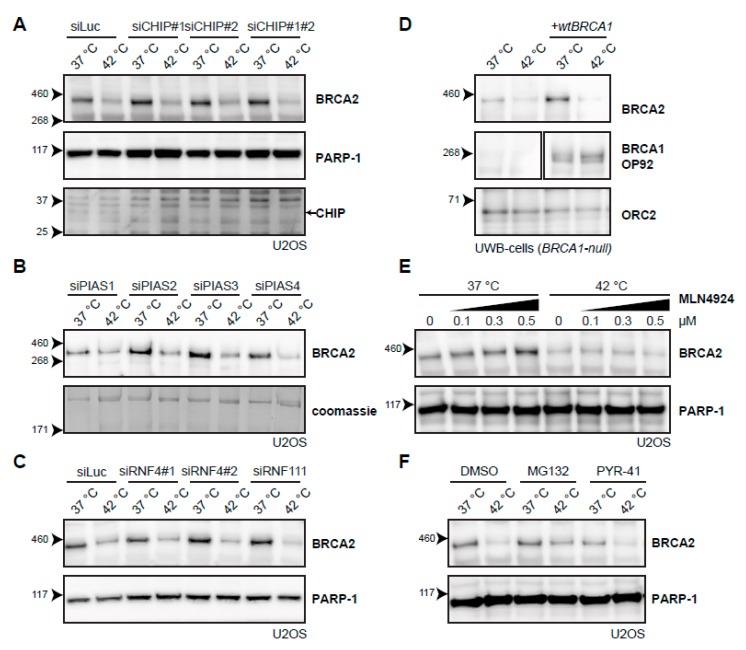
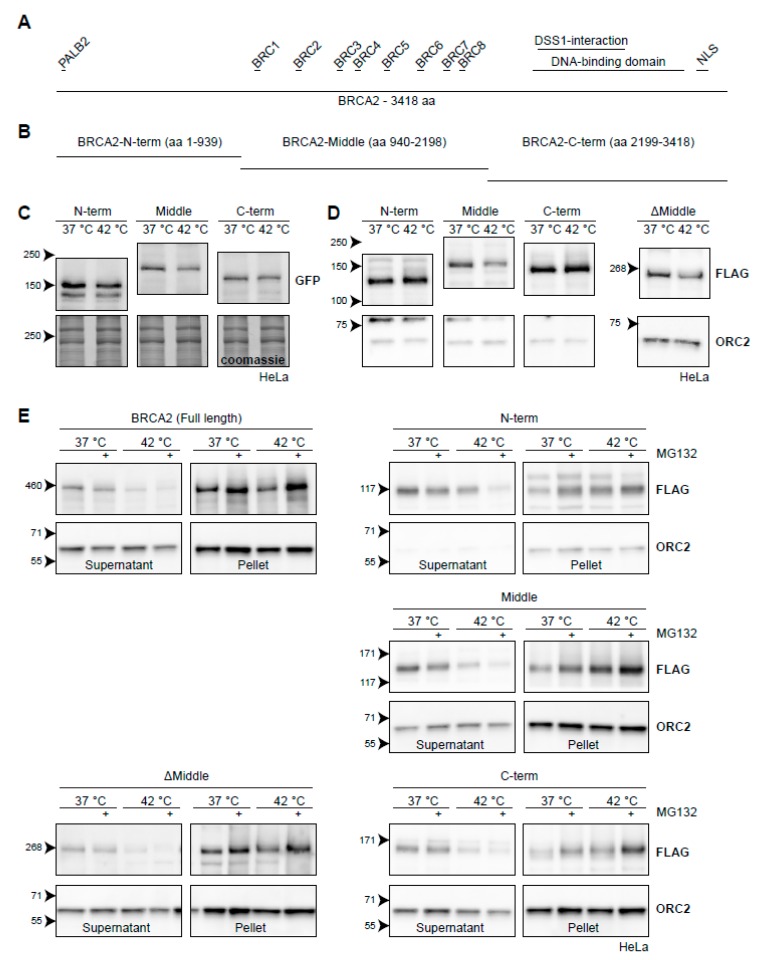
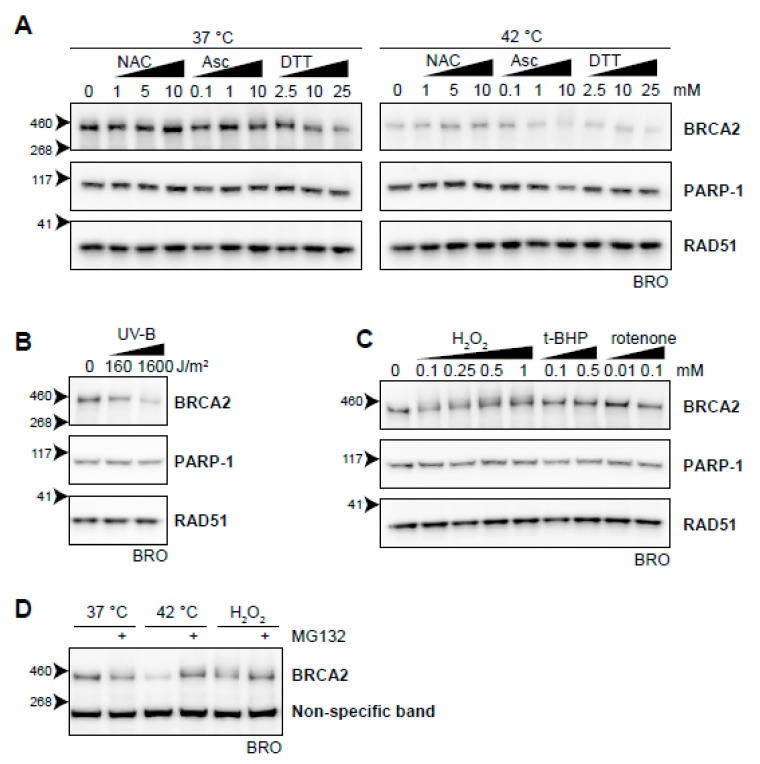
The DNA damage response (DDR) is a designation for a number of pathways that protects our DNA from various damaging agents. In normal cells, the DDR is extremely important for maintaining genome integrity, but in cancer cells these mechanisms counteract therapy-induced DNA damage. Inhibition of the DDR could therefore be used to increase the efficacy of anti-cancer treatments. Hyperthermia is an example of such a treatment-it inhibits a sub-pathway of the DDR, called homologous recombination (HR). It does so by inducing proteasomal degradation of BRCA2 -one of the key HR factors. Understanding the precise mechanism that mediates this degradation is important for our understanding of how hyperthermia affects therapy and how homologous recombination and BRCA2 itself function. In addition, mechanistic insight into the process of hyperthermia-induced BRCA2 degradation can yield new therapeutic strategies to enhance the effects of local hyperthermia or to inhibit HR. Here, we investigate the mechanisms driving hyperthermia-induced BRCA2 degradation. We find that BRCA2 degradation is evolutionarily conserved, that BRCA2 stability is dependent on HSP90, that ubiquitin might not be involved in directly targeting BRCA2 for protein degradation via the proteasome, and that BRCA2 degradation might be modulated by oxidative stress and radical scavengers.
Keywords: BRCA2; HSP90; RAD51; SILAC mass spectrometry; homologous recombination; hyperthermia; protein degradation; reactive oxygen species; ubiquitin.
Conflict of interest statement
The authors declare no conflict of interest
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
