

Gene duplication and genetic innovation in cereal genomes
- PMID: 30651279
- PMCID: PMC6360818
- DOI: 10.1101/gr.237511.118
Gene duplication and genetic innovation in cereal genomes
Abstract
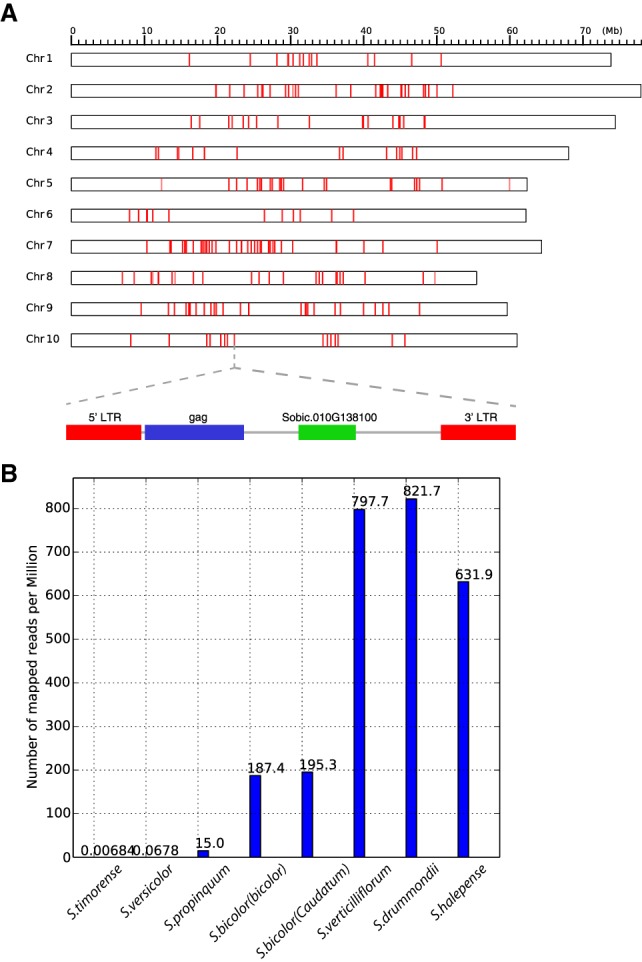
Organisms continuously require genetic variation to adapt to fluctuating environments, yet major evolutionary events are episodic, making the relationship between genome evolution and organismal adaptation of considerable interest. Here, by genome-wide comparison of sorghum, maize, and rice SNPs, we investigated reservoirs of genetic variations with high precision. For sorghum and rice, which have not experienced whole-genome duplication in 96 million years or more, tandem duplicates accumulate relatively more SNPs than paralogous genes retained from genome duplication. However, maize, which experienced lineage-specific genome duplication and has a relatively larger supply of paralogous duplicates, shows SNP enrichment in paralogous genes. The proportion of genes showing signatures of recent positive selection is higher in small-scale (tandem and transposed) than genome-scale duplicates in sorghum, but the opposite is true in maize. A large proportion of recent duplications in rice are species-specific; however, most recent duplications in sorghum are derived from ancestral gene families. A new retrotransposon family was also a source of many recent sorghum duplications, illustrating a role in providing variation for genetic innovations. This study shows that diverse evolutionary mechanisms provide the raw genetic material for adaptation in taxa with divergent histories of genome evolution.
© 2019 Guo et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





References
-
- Brar DS, Singh K. 2011. Oryza. In Wild crop relatives: genomic and breeding resources: cereals (ed. Kole C), pp. 321–367. Springer, Heidelberg, Germany.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources