

Superficial Layers Suppress the Deep Layers to Fine-tune Cortical Coding
- PMID: 30651326
- PMCID: PMC6507093
- DOI: 10.1523/JNEUROSCI.1459-18.2018
Superficial Layers Suppress the Deep Layers to Fine-tune Cortical Coding
Abstract
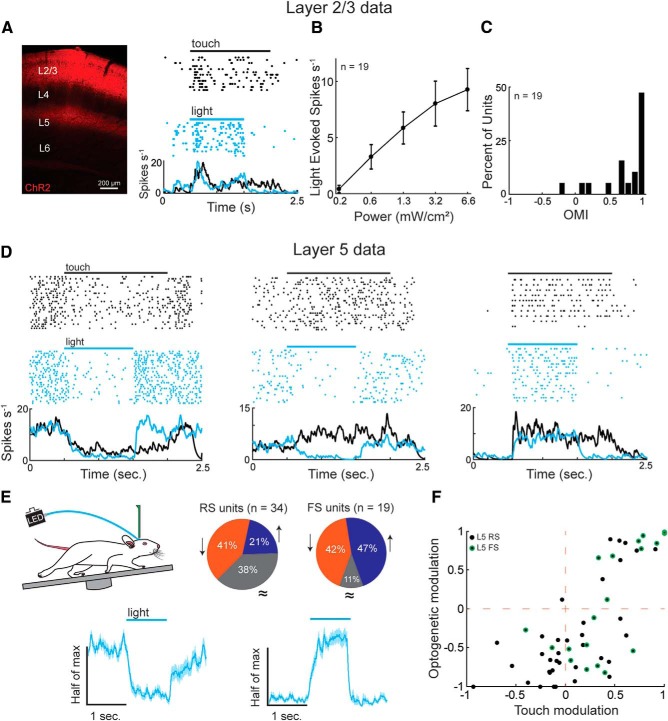
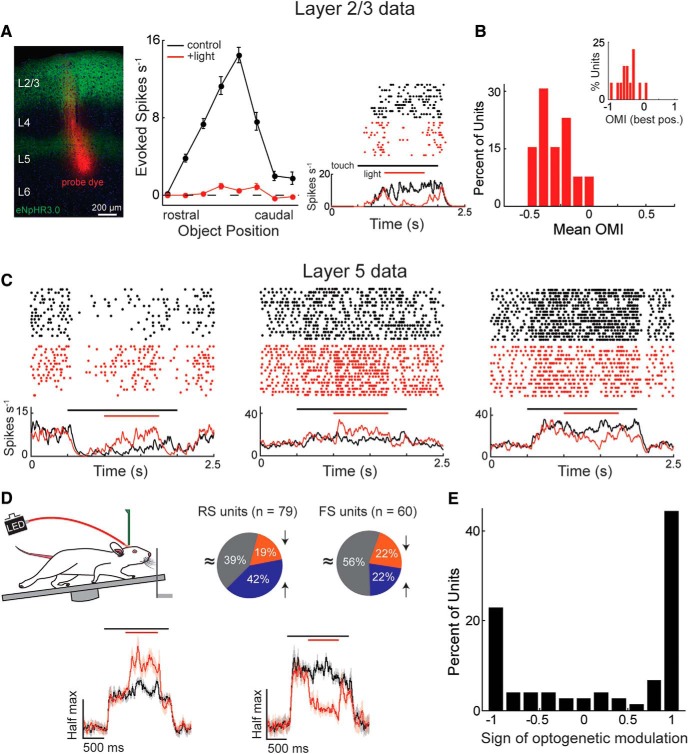
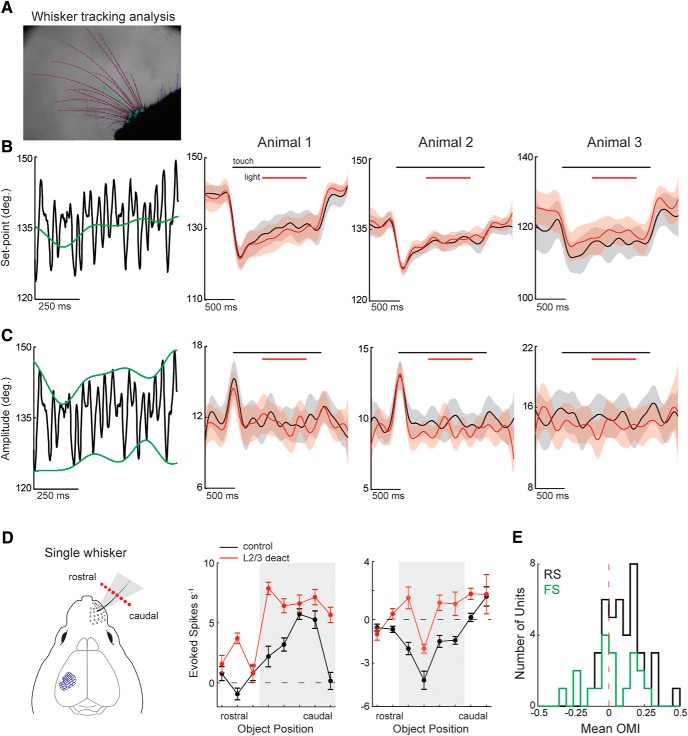
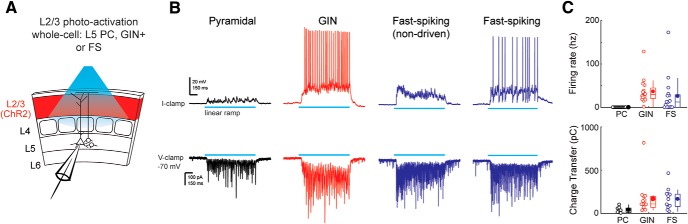
The descending microcircuit from layer 2/3 (L2/3) to layer 5 (L5) is one of the strongest excitatory pathways in the cortex, presumably forming a core component of its feedforward hierarchy. To date, however, no experiments have selectively tested the impact of L2/3 activity on L5 during active sensation. We used optogenetic, cell-type-specific manipulation of L2/3 neurons in the barrel cortex of actively sensing mice (of either sex) to elucidate the significance of this pathway to sensory coding in L5. Contrary to standard models, activating L2/3 predominantly suppressed spontaneous activity in L5, whereas deactivating L2/3 mainly facilitated touch responses in L5. Somatostatin interneurons are likely important to this suppression because their optogenetic deactivation significantly altered the functional impact of L2/3 onto L5. The net effect of L2/3 was to enhance the stimulus selectivity and expand the range of L5 output. These data imply that the core cortical pathway increases the selectivity and expands the range of cortical output through feedforward inhibition.SIGNIFICANCE STATEMENT The primary sensory cortex contains six distinct layers that interact to form the basis of our perception. While rudimentary patterns of connectivity between the layers have been outlined quite extensively in vitro, functional relationships in vivo, particularly during active sensation, remain poorly understood. We used cell-type-specific optogenetics to test the functional relationship between layer 2/3 and layer 5. Surprisingly, we discovered that L2/3 primarily suppresses cortical output from L5. The recruitment of somatostatin-positive interneurons is likely fundamental to this relationship. The net effect of this translaminar suppression is to enhance the selectivity and expand the range of receptive fields, therefore potentially sharpening the perception of space.
Keywords: active sensing; circuits; cortex; receptive fields; sensory coding; translaminar.
Copyright © 2019 the authors 0270-6474/19/392052-13$15.00/0.
Figures
Similar articles
-
Translaminar recurrence from layer 5 suppresses superficial cortical layers.Nat Commun. 2022 May 11;13(1):2585. doi: 10.1038/s41467-022-30349-w. Nat Commun. 2022. PMID: 35546553 Free PMC article.
-
Diversity and Connectivity of Layer 5 Somatostatin-Expressing Interneurons in the Mouse Barrel Cortex.J Neurosci. 2018 Feb 14;38(7):1622-1633. doi: 10.1523/JNEUROSCI.2415-17.2017. Epub 2018 Jan 11. J Neurosci. 2018. PMID: 29326172 Free PMC article.
-
A direct translaminar inhibitory circuit tunes cortical output.Nat Neurosci. 2015 Nov;18(11):1631-40. doi: 10.1038/nn.4123. Epub 2015 Sep 28. Nat Neurosci. 2015. PMID: 26414615 Free PMC article.
-
Thalamocortical control of feed-forward inhibition in awake somatosensory 'barrel' cortex.Philos Trans R Soc Lond B Biol Sci. 2002 Dec 29;357(1428):1717-27. doi: 10.1098/rstb.2002.1156. Philos Trans R Soc Lond B Biol Sci. 2002. PMID: 12626006 Free PMC article. Review.
-
Inhibitory Circuits in Cortical Layer 5.Front Neural Circuits. 2016 May 6;10:35. doi: 10.3389/fncir.2016.00035. eCollection 2016. Front Neural Circuits. 2016. PMID: 27199675 Free PMC article. Review.
Cited by
-
Optogenetic stimulation shapes dendritic trees of infragranular cortical pyramidal cells.Front Cell Neurosci. 2023 Aug 1;17:1212483. doi: 10.3389/fncel.2023.1212483. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37587917 Free PMC article.
-
Effects of optogenetic silencing the anterior cingulate cortex in a delayed non-match to trajectory task.Oxf Open Neurosci. 2024 Feb 8;3:kvae002. doi: 10.1093/oons/kvae002. eCollection 2024. Oxf Open Neurosci. 2024. PMID: 38595941 Free PMC article.
-
Touch-evoked traveling waves establish a translaminar spacetime code.Sci Adv. 2025 Jan 31;11(5):eadr4038. doi: 10.1126/sciadv.adr4038. Epub 2025 Jan 31. Sci Adv. 2025. PMID: 39889002 Free PMC article.
-
Thalamocortical Mechanisms Regulating the Relationship between Transient Beta Events and Human Tactile Perception.Cereb Cortex. 2022 Feb 8;32(4):668-688. doi: 10.1093/cercor/bhab221. Cereb Cortex. 2022. PMID: 34401898 Free PMC article.
-
Sensorimotor strategies and neuronal representations for shape discrimination.Neuron. 2021 Jul 21;109(14):2308-2325.e10. doi: 10.1016/j.neuron.2021.05.019. Epub 2021 Jun 15. Neuron. 2021. PMID: 34133944 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases