

Heterochromatin suppresses gross chromosomal rearrangements at centromeres by repressing Tfs1/TFIIS-dependent transcription
- PMID: 30652128
- PMCID: PMC6329695
- DOI: 10.1038/s42003-018-0251-z
Heterochromatin suppresses gross chromosomal rearrangements at centromeres by repressing Tfs1/TFIIS-dependent transcription
Abstract
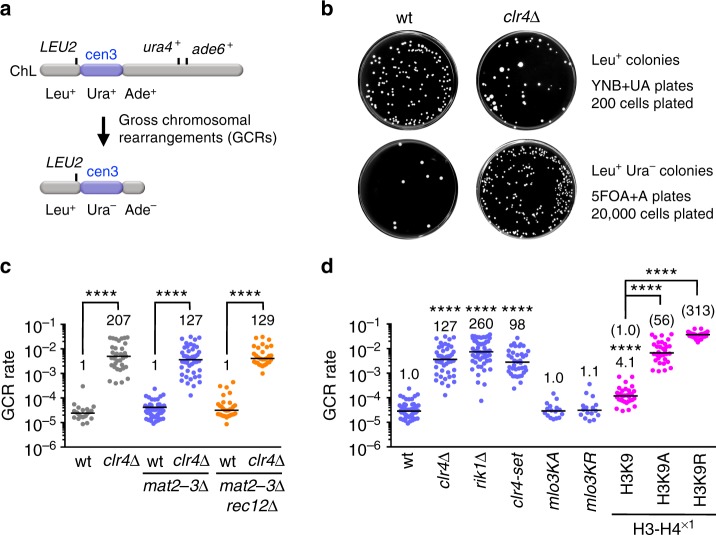
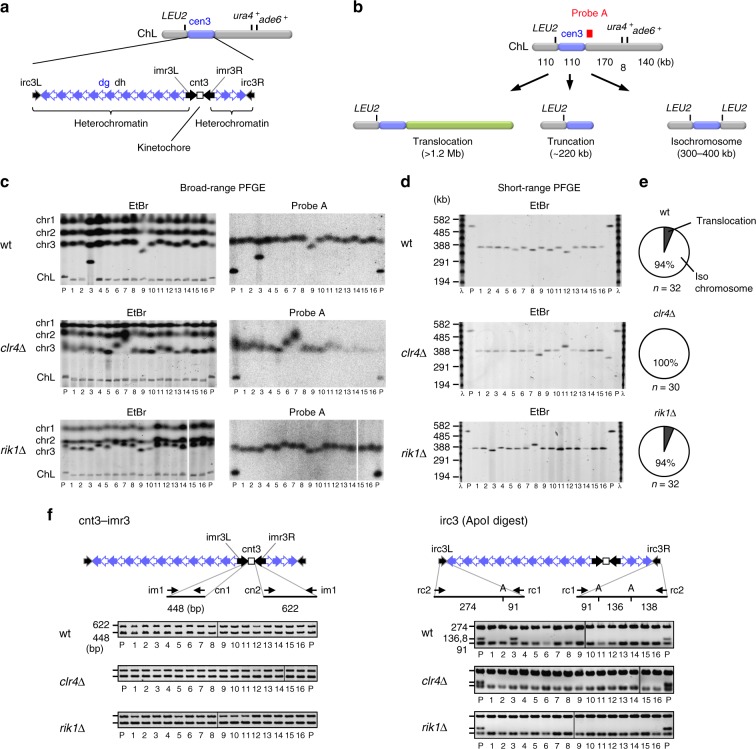
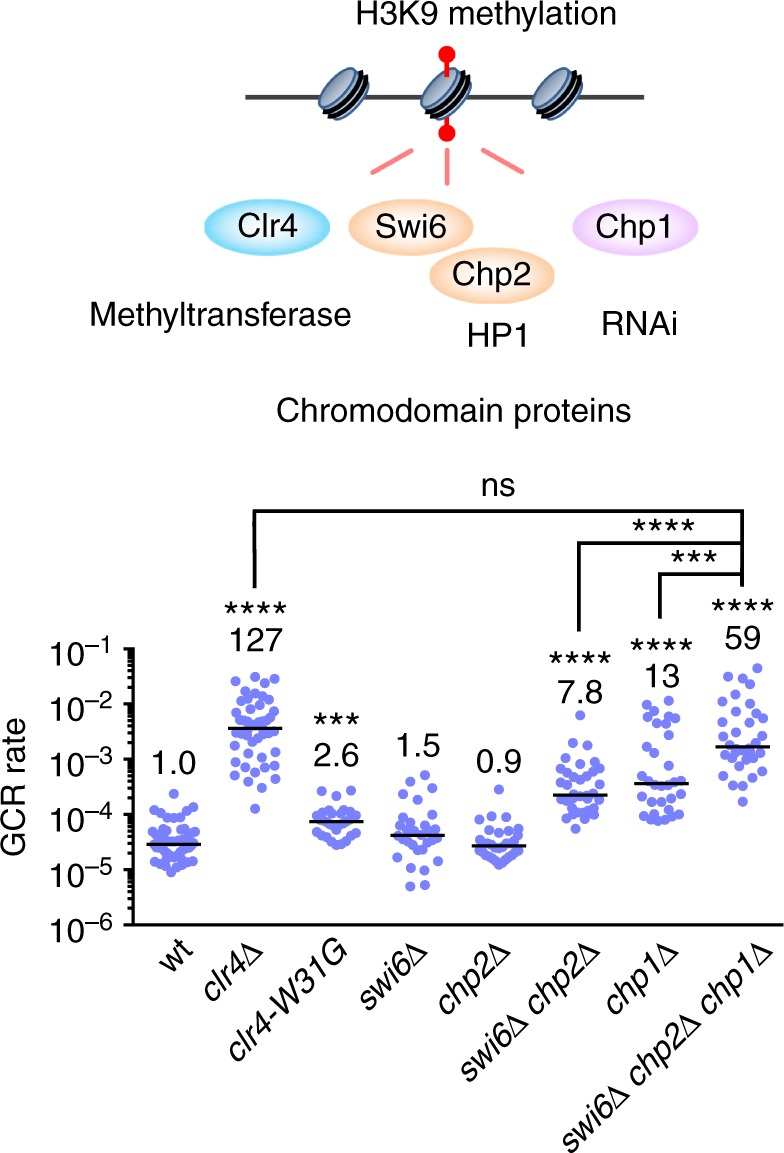
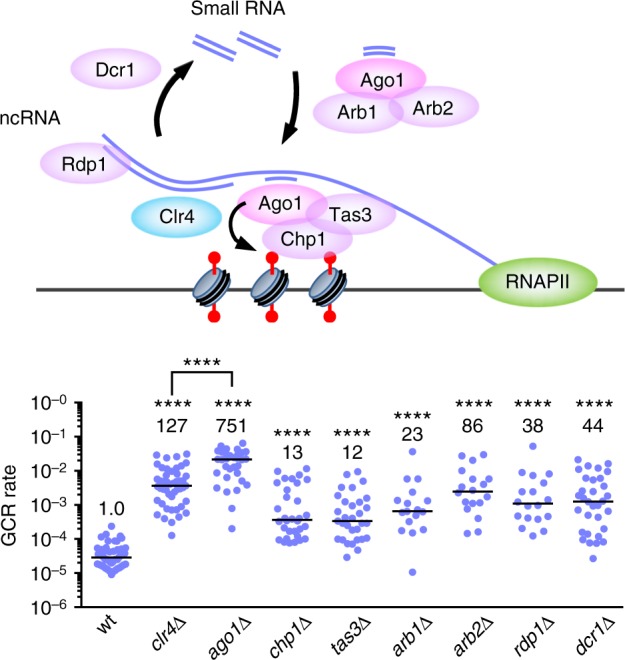
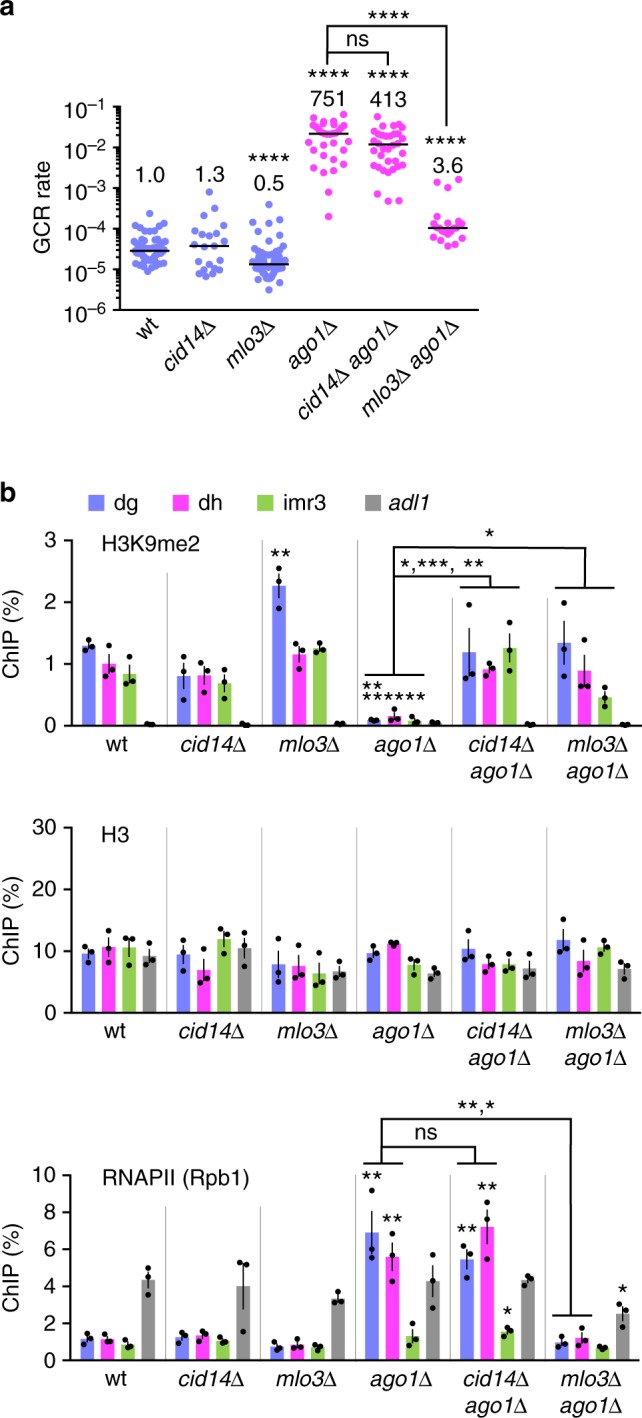
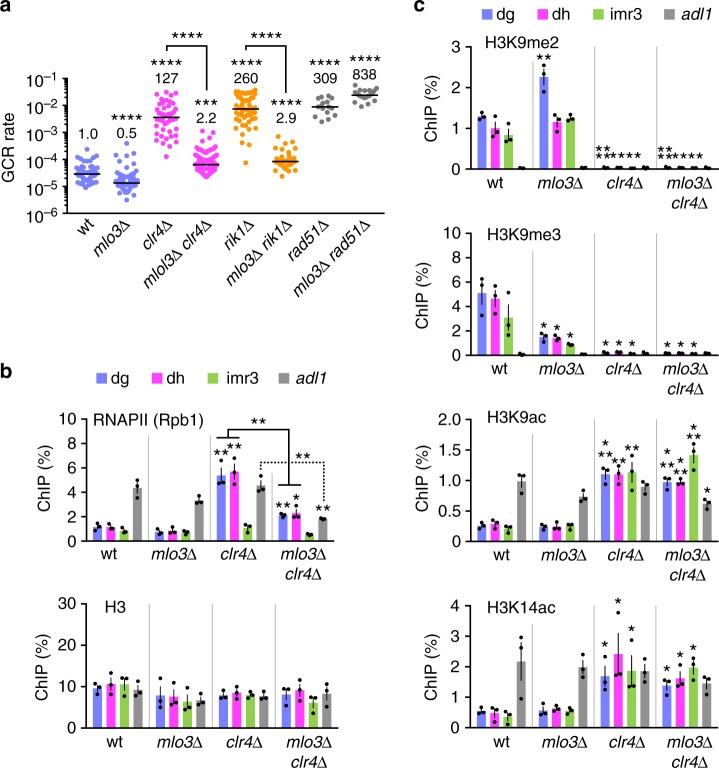
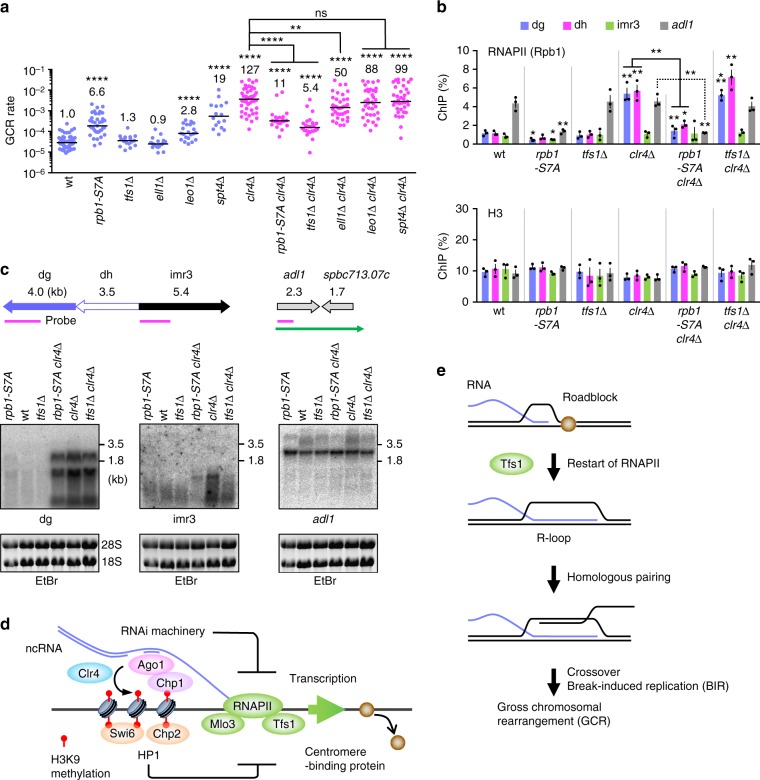
Heterochromatin, characterized by histone H3 lysine 9 (H3K9) methylation, assembles on repetitive regions including centromeres. Although centromeric heterochromatin is important for correct segregation of chromosomes, its exact role in maintaining centromere integrity remains elusive. Here, we found in fission yeast that heterochromatin suppresses gross chromosomal rearrangements (GCRs) at centromeres. Mutations in Clr4/Suv39 methyltransferase increased the formation of isochromosomes, whose breakpoints were located in centromere repeats. H3K9A and H3K9R mutations also increased GCRs, suggesting that Clr4 suppresses centromeric GCRs via H3K9 methylation. HP1 homologs Swi6 and Chp2 and the RNAi component Chp1 were the chromodomain proteins essential for full suppression of GCRs. Remarkably, mutations in RNA polymerase II (RNAPII) or Tfs1/TFIIS, the transcription factor that facilitates restart of RNAPII after backtracking, specifically bypassed the requirement of Clr4 for suppressing GCRs. These results demonstrate that heterochromatin suppresses GCRs by repressing Tfs1-dependent transcription of centromere repeats.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
