

Regulation of RNA editing by RNA-binding proteins in human cells
- PMID: 30652130
- PMCID: PMC6331435
- DOI: 10.1038/s42003-018-0271-8
Regulation of RNA editing by RNA-binding proteins in human cells
Abstract
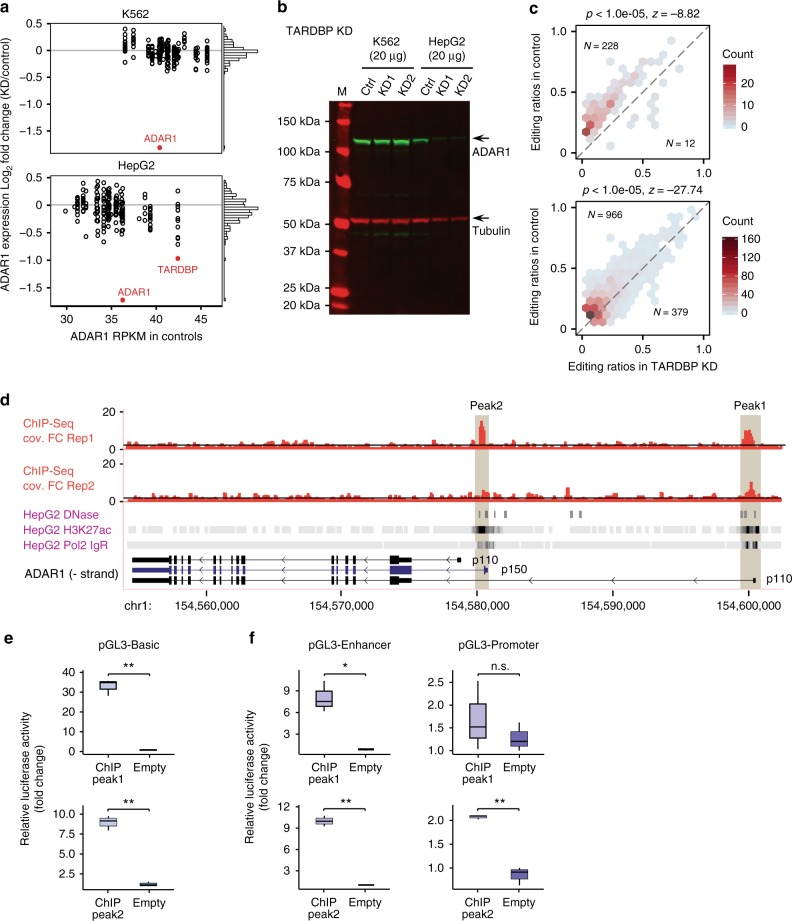
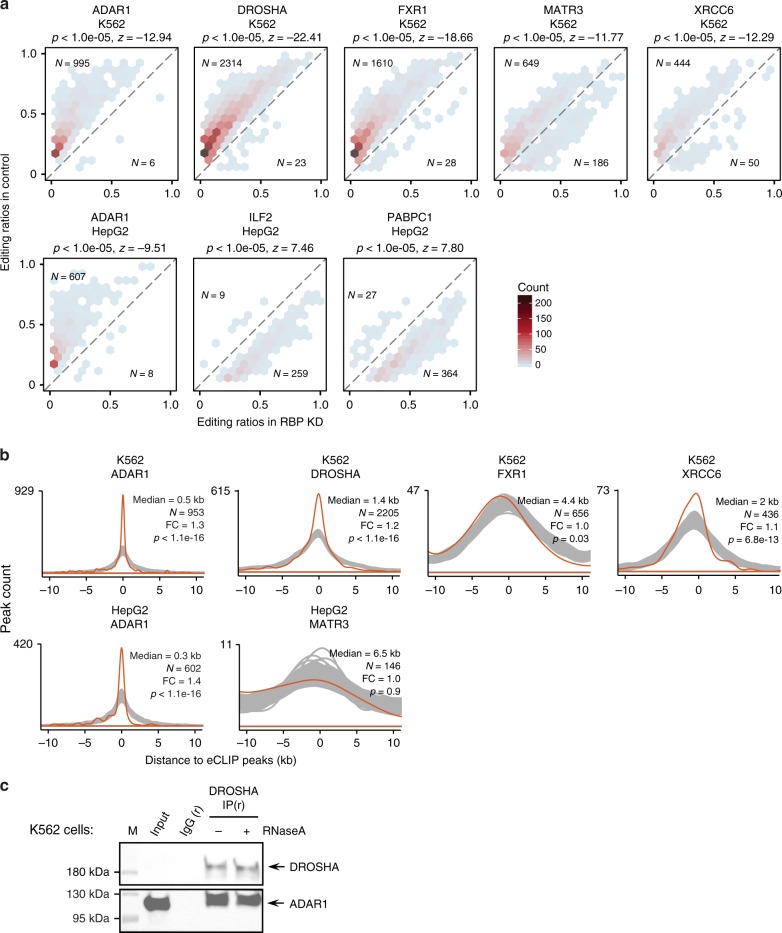
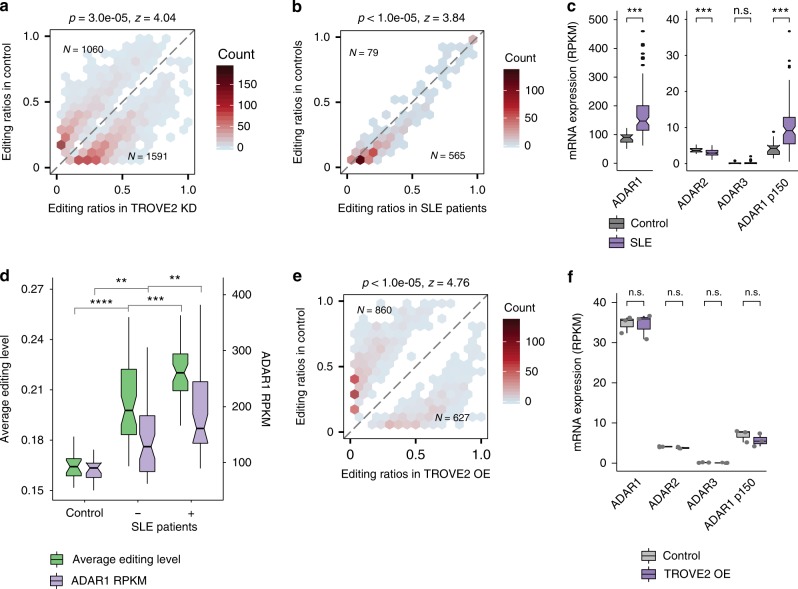
Adenosine-to-inosine (A-to-I) editing, mediated by the ADAR enzymes, diversifies the transcriptome by altering RNA sequences. Recent studies reported global changes in RNA editing in disease and development. Such widespread editing variations necessitate an improved understanding of the regulatory mechanisms of RNA editing. Here, we study the roles of >200 RNA-binding proteins (RBPs) in mediating RNA editing in two human cell lines. Using RNA-sequencing and global protein-RNA binding data, we identify a number of RBPs as key regulators of A-to-I editing. These RBPs, such as TDP-43, DROSHA, NF45/90 and Ro60, mediate editing through various mechanisms including regulation of ADAR1 expression, interaction with ADAR1, and binding to Alu elements. We highlight that editing regulation by Ro60 is consistent with the global up-regulation of RNA editing in systemic lupus erythematosus. Additionally, most key editing regulators act in a cell type-specific manner. Together, our work provides insights for the regulatory mechanisms of RNA editing.
Conflict of interest statement
G.W.Y. is a co-founder of Locana and Eclipse Bioinnovations and member of the scientific advisory boards of Locana, Eclipse Bioinnovations and Aquinnah Pharmaceuticals. E.V.N is a co-founder and member of the scientific advisory board of Eclipse BioInnovations. The terms of these arrangements have been reviewed and approved by the University of California, San Diego in accordance with its conflict of interest policies. The remaining authors declare no competing interests.
Figures



Similar articles
-
Adenosine-to-Inosine RNA Editing Enzyme ADAR and microRNAs.Methods Mol Biol. 2021;2181:83-95. doi: 10.1007/978-1-0716-0787-9_6. Methods Mol Biol. 2021. PMID: 32729076
-
Adenosine-to-inosine Alu RNA editing controls the stability of the pro-inflammatory long noncoding RNA NEAT1 in atherosclerotic cardiovascular disease.J Mol Cell Cardiol. 2021 Nov;160:111-120. doi: 10.1016/j.yjmcc.2021.07.005. Epub 2021 Jul 21. J Mol Cell Cardiol. 2021. PMID: 34302813 Free PMC article.
-
Combinatory RNA-Sequencing Analyses Reveal a Dual Mode of Gene Regulation by ADAR1 in Gastric Cancer.Dig Dis Sci. 2018 Jul;63(7):1835-1850. doi: 10.1007/s10620-018-5081-9. Epub 2018 Apr 25. Dig Dis Sci. 2018. PMID: 29691780
-
In cancer, A-to-I RNA editing can be the driver, the passenger, or the mechanic.Drug Resist Updat. 2017 May;32:16-22. doi: 10.1016/j.drup.2017.09.001. Epub 2017 Oct 4. Drug Resist Updat. 2017. PMID: 29145975 Review.
-
A-to-I RNA editing and hematopoiesis.Exp Hematol. 2024 Nov;139:104621. doi: 10.1016/j.exphem.2024.104621. Epub 2024 Aug 24. Exp Hematol. 2024. PMID: 39187172 Review.
Cited by
-
Ro60-Roles in RNA Processing, Inflammation, and Rheumatic Autoimmune Diseases.Int J Mol Sci. 2024 Jul 14;25(14):7705. doi: 10.3390/ijms25147705. Int J Mol Sci. 2024. PMID: 39062948 Free PMC article. Review.
-
LoDEI: a robust and sensitive tool to detect transcriptome-wide differential A-to-I editing in RNA-seq data.Nat Commun. 2024 Oct 23;15(1):9121. doi: 10.1038/s41467-024-53298-y. Nat Commun. 2024. PMID: 39443485 Free PMC article.
-
Cutting Edge: Reduced Adenosine-to-Inosine Editing of Endogenous Alu RNAs in Severe COVID-19 Disease.J Immunol. 2021 Apr 15;206(8):1691-1696. doi: 10.4049/jimmunol.2001428. Epub 2021 Mar 29. J Immunol. 2021. PMID: 33782089 Free PMC article.
-
Role of RNA modifications in blood development and regeneration.Exp Hematol. 2024 Oct;138:104279. doi: 10.1016/j.exphem.2024.104279. Epub 2024 Jul 14. Exp Hematol. 2024. PMID: 39009277 Review.
-
Establishment of chemically oligomerizable TAR DNA-binding protein-43 which mimics amyotrophic lateral sclerosis pathology in mammalian cells.Lab Invest. 2021 Oct;101(10):1331-1340. doi: 10.1038/s41374-021-00623-4. Epub 2021 Jun 15. Lab Invest. 2021. PMID: 34131277
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
