

Single-molecule dynamics and genome-wide transcriptomics reveal that NF-kB (p65)-DNA binding times can be decoupled from transcriptional activation
- PMID: 30653501
- PMCID: PMC6353211
- DOI: 10.1371/journal.pgen.1007891
Single-molecule dynamics and genome-wide transcriptomics reveal that NF-kB (p65)-DNA binding times can be decoupled from transcriptional activation
Abstract
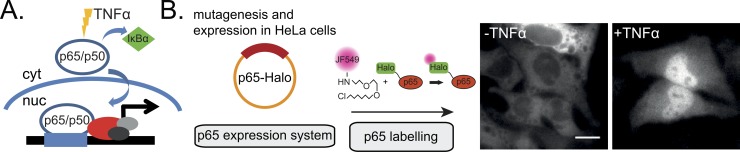
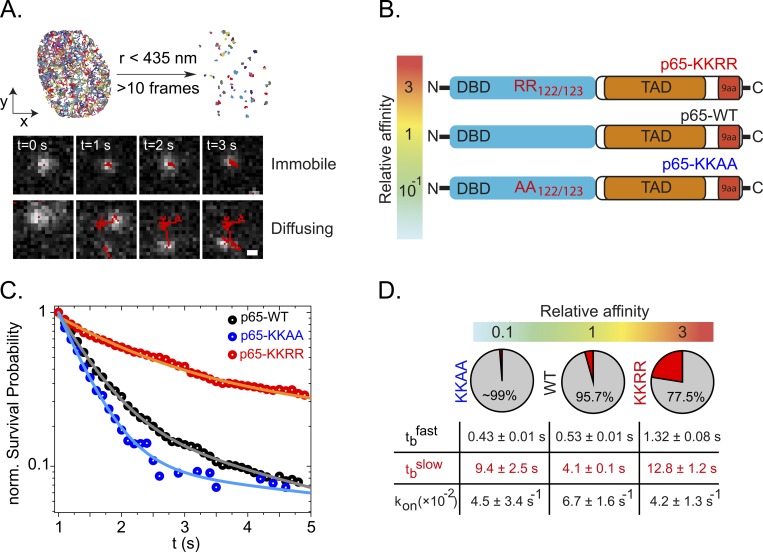
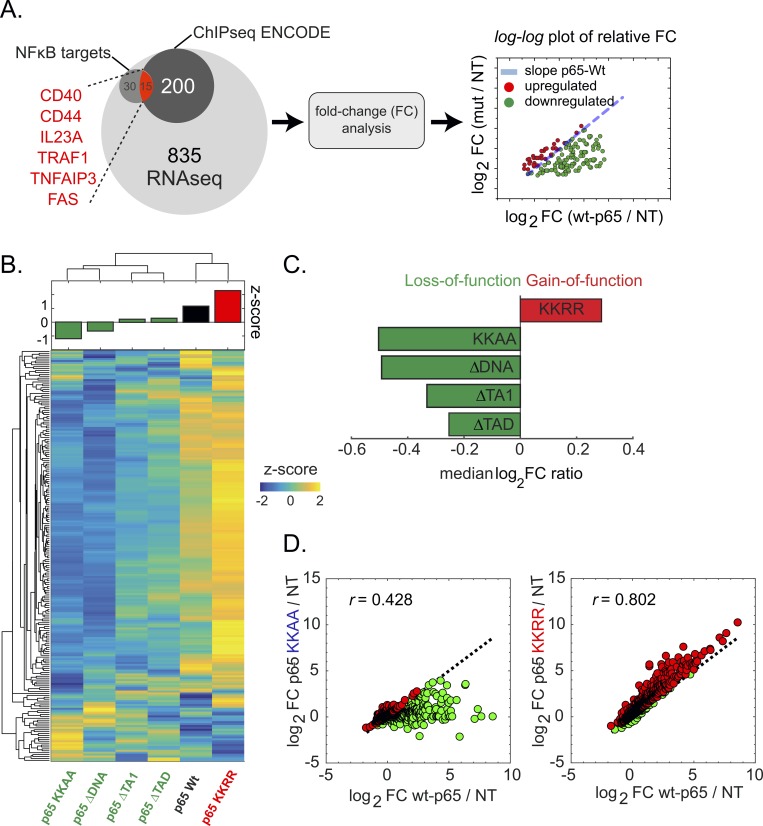
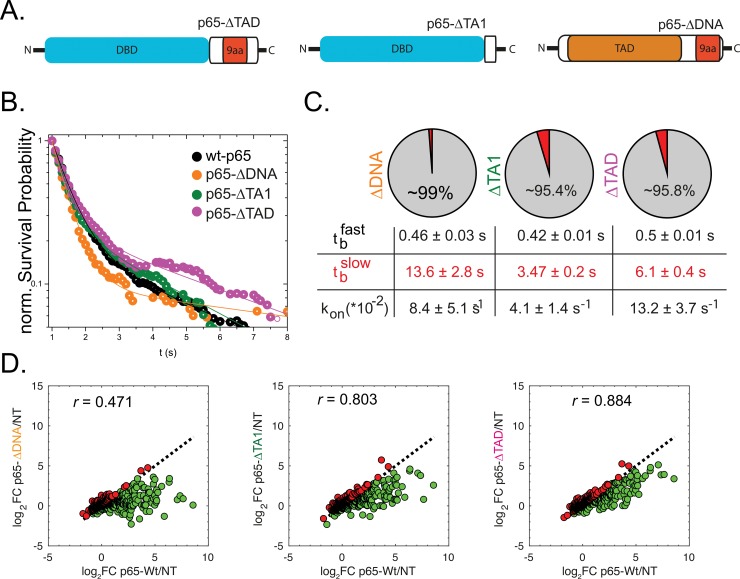
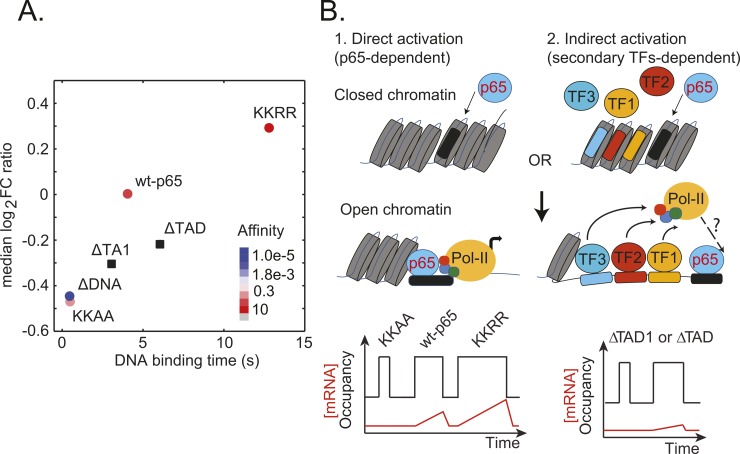
Transcription factors (TFs) regulate gene expression in both prokaryotes and eukaryotes by recognizing and binding to specific DNA promoter sequences. In higher eukaryotes, it remains unclear how the duration of TF binding to DNA relates to downstream transcriptional output. Here, we address this question for the transcriptional activator NF-κB (p65), by live-cell single molecule imaging of TF-DNA binding kinetics and genome-wide quantification of p65-mediated transcription. We used mutants of p65, perturbing either the DNA binding domain (DBD) or the protein-protein transactivation domain (TAD). We found that p65-DNA binding time was predominantly determined by its DBD and directly correlated with its transcriptional output as long as the TAD is intact. Surprisingly, mutation or deletion of the TAD did not modify p65-DNA binding stability, suggesting that the p65 TAD generally contributes neither to the assembly of an "enhanceosome," nor to the active removal of p65 from putative specific binding sites. However, TAD removal did reduce p65-mediated transcriptional activation, indicating that protein-protein interactions act to translate the long-lived p65-DNA binding into productive transcription.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Baeuerle PA, Baltimore D. IκB: A specific inhibitor of the NF-κB transcription factor. Science (80-). 1988;242: 540–546. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
